









 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
España es el primer productor de porcino en Europa. Los 30 millones de cerdos cebados en 2017 equivalen al 20% de la producción europea (Unión Europea, 2018). Las deyecciones ganaderas generadas, mayoritariamente en forma de purines, se usan como fertilizante para la producción de grano y forraje, siendo la aplicación directa en campo la más común por ser la opción más económica. Las dosis aplicadas suelen basarse en la demanda de N de los cultivos. El uso de purín como fertilizante permite al agricultor acoplarse a las exigencias de la Política Agrícola Común establecida en la Unión Europea (2013) que incluye, entre otros, el mantenimiento de la materia orgánica (MO) y la estructura del suelo, y el evitar el deterioro de los hábitats. Sin embargo, debido a la gran disponibilidad de purín y la necesidad de vaciar las balsas de almacenamiento mediante aplicaciones en las fincas cercanas, no se puede descartar que en determinadas situaciones pueda existir una sobrefertilización. Los purines, además de aportar macro y micronutrientes, suelen incluir insumos de alimentación y sanidad animal (aditivos y productos zoosanitarios y farmacológicos) y sus derivados metabólicos y de degradación (De la Torre, Jiménez, Carballo Fernandez, Roset & Muñoz, 2000), por lo que las aplicaciones excesivas al suelo constituyen un riesgo ambiental. Adicionalmente, Murchie, Blackshaw, Gordon & Christie (2015) mencionan que son de particular preocupación las acumulaciones de metales pesados.
Los purines de cerdo, según el tipo de granja de la que provienen: cerdos en engorde (PE), madres reproductoras (PM) o en ciclo cerrado presentan composiciones diferentes (Sánchez & González, 2005; Yagüe, Bosch-Serra & Boixadera, 2012). Por ello, las aplicaciones de PE y PM a largo plazo podrían incidir sobre las propiedades del suelo de manera diferenciada, cambios a los que podrían ser sensibles determinados organismos del suelo, como las lombrices de tierra (Renaud et al., 2017).
Las lombrices de tierra juegan un papel importante en la regulación de los servicios ecosistémicos del suelo que dan soporte a la agricultura, principalmente en la descomposición de la MO, ciclo de nutrientes y actividades de bioturbación como la mezcla de materiales y formación de bioporos. Según Manono (2016), las lombrices de tierra son muy sensibles a prácticas de manejo del suelo afectándoles a nivel taxonómico (diversidad de especies), de grupos funcionales y globalmente (abundancia y biomasa). Así, las lombrices de tierra se consideran bioindicadoras de impactos ambientales en entornos rurales asociados al laboreo o a la contaminación del suelo (Paoletti, 1999). El tipo y la cantidad de fertilizante orgánico aplicado también puede afectar a la comunidad de lombrices de tierra (Koblenz, Tischer, Rücknagel & Christen, 2015; Murchie et al., 2015), por lo que la evaluación de los impactos sobre esta comunidad tras un uso continuado resulta ser de interés. En estudios previos, principalmente en pastizales, se ha observado que los purines: (i) tienen efectos tóxicos transitorios (por su contenido en N amoniacal y otras sales) sobre las lombrices de tierra de hábito superficial (Curry, 1976), (ii) tienen un efecto positivo pero inferior al del estiércol debido a su menor contenido en MO (Timmerman, Bos, Ouwehand & Goede, 2006), (iii) con aplicaciones a dosis elevadas, podrían reducir la biomasa y abundancia de lombrices de tierra (Edwards & Lofty, 1982; Hansen & Engelstad, 1999; Paoletti, 1999), (iv) impactan de manera diferencial según el método de aplicación (superficial o inyección) utilizado (De Goede, Brussaard & Akkermans, 2003), (v) aplicados en dosis crecientes (tanto de bovino como de porcino) afectan de manera distinta a las comunidades de lombrices de tierra (Koblenz et al., 2015; Murchie et al., 2015), (vi) pueden afectar al crecimiento de la población de lombrices de tierra y éste puede estar limitado por el enriquecimiento de Cu y Zn (Murchie et al., 2015).
Por lo tanto, el estudio de distintos parámetros asociados a la presencia de lombrices de tierra podría ser interesante para detectar zonas donde la calidad del suelo esté en riesgo debido a la sobrefertilización. Este estudio se desarrolló bajo un ambiente Mediterráneo semiárido, en un sistema cerealístico de agricultura de secano y en una campaña en barbecho. El objetivo fue evaluar el efecto de la fertilización precedente (mantenida durante 12 años) con purín porcino de diferentes orígenes (PE y PM) sobre la población de lombrices de tierra, tanto a nivel taxonómico (composición de especies) como global (abundancia y biomasa).
La hipótesis de esta investigación es que la abundancia, biomasa y diversidad de especies de lombrices de tierra podrían ser bioindicadoras de prácticas de sobrefertilización con purines y que, además, la afectación sería diferente según el origen del purín porcino (cerdo de engorde o de madres reproductoras).
MATERIALES Y MÉTODOS
El estudio se ubicó en una finca de secano del NE de España, en una zona de clima Mediterráneo semiárido (MAPA, 1989) con valores medios anuales de 370 mm en precipitación y de 13ºC en temperatura. Las coordenadas de la finca son 41º52'29''N, 1º09'10''E y la altitud es de 443 m s. n. m. El suelo se clasifica como Typic Xerofluvent (Soil Survey Staff, 2014). El horizonte superficial (0-0,30 m) presenta un pH (potenciometría, 1:2,5; suelo: agua destilada) de 8,2; una conductividad eléctrica (1:5; suelo: agua destilada, 25°C) de 0,18 dS m−1; un contenido de materia orgánica medio de 20,1 g kg−1 (método de Walkley y Black) y de carbonato de calcio equivalente de 300 g kg−1 (método de calcímetro de Bernard). La textura (clasificación USDA) es franco limosa: 131 g kg−1 arena, 609 g kg−1 limo y 260 g kg−1 arcilla (método de la pipeta).
En la finca experimental se estableció una rotación de cereales de invierno: cebada (Hordeum vulgare L.) y trigo (Triticum aestivum L.), con algún año en barbecho como en el año del presente estudio (campaña 2016−2017). La paja se recolecta y empaca tras la cosecha (julio), y el rastrojo se entierra a final del verano (septiembre) mediante laboreo con rastras de discos (~0,15 m). Las diferentes estrategias de abonado previas a este estudio incluyeron, durante doce años, purín de cerdo de engorde (PE), purín de madres reproductoras (PM) y una estrategia con fertilización mineral (FM). El presente estudio se centra en siete tratamientos (se incluyen las equivalencias en cantidad de N aplicado durante la campaña 2015−16): un tratamiento con fertilización mineral que actúa como control (0−FM), en donde el N se aplica todo en cobertera (macollaje de cereales 21−24 de la escala decimal Zadoks, Chang & Konzak (1974)) como nitrato amónico cálcico; tres dosis de PE (en peso fresco) que fueron de 30 t ha−1 año−1 (30PE−0 aplicado en pre-siembra), 40 y 60 t ha−1 año−1 (0−40PE y 0−60PE respectivamente, aplicados en cobertera), dos dosis de PM (en peso fresco) que fueron de 60 y 90 t ha−1 año−1 (0−60PM y 0−90PM respectivamente, aplicados en cobertera) y un tratamiento donde se combinó la aplicación de PE en pre-siembra a dosis de 30 t ha−1 año−1 con PM en cobertera a dosis de 90 t ha−1 año−1 (30PE−90PM). Las equivalencias de las dosis en cantidad de N aplicado durante la campaña 2015−2016, previa al muestreo, se describen en la Tabla 1. En esta campaña las producciones (grano sobre materia seca) oscilaron entre 5217 kg ha-1 (FM) y 6062 kg ha-1 (30PE−90PM) con una producción media de todos los tratamientos con purines de 5730 kg ha-1 (±314, desviación estándar). Los purines se aplicaron mediante el método habitual de placa de salpicadura (en abanico).
El diseño experimental consistió en bloques completos de parcelas al azar, con tres repeticiones para cada tratamiento de fertilización. Las parcelas tenían una superficie de 274 m2 (11 m de ancho y 25 m de largo) excepto el tratamiento mineral que contaba con una superficie de 174 m2 (7 m de ancho y 25 m de largo).
El muestreo de lombrices de tierra se realizó el 16 de marzo de 2017, tras 13 y 17 meses después de la última aplicación de fertilizante en cobertera y pre-siembra, respectivamente. El último laboreo del suelo se realizó con un pase de grada de discos en octubre de 2016 para el control de las malas hierbas. La elección de la época de muestreo, final de invierno e inicio de primavera, se basó en las recomendaciones de Pérès et al. (2014), por considerarse el periodo activo de esta población. El contenido medio de agua en el suelo se cuantificó en las muestras y fue igual al 16% (p/p, humedad gravimétrica). Las lombrices de tierra se extrajeron siguiendo el protocolo de muestreo de la norma española UNE-ISO 23611-1 (AENOR, 2009). El procedimiento técnico consistió en la excavación de monolitos de suelo (0,25 m x 0,25 m) hasta una profundidad de 0,20 m. Las lombrices se recogieron manualmente, se lavaron con agua y conservaron inmediatamente en etanol (70%) previa la fijación con formaldehído (AENOR, 2009). Se obtuvieron un total de 21 muestras de suelo.
Tabla 1 Nitrógeno aplicado en la campaña de cereal previa a la del muestreo de lombrices (campaña 2015−2016) en los diferentes tratamientos de fertilización.
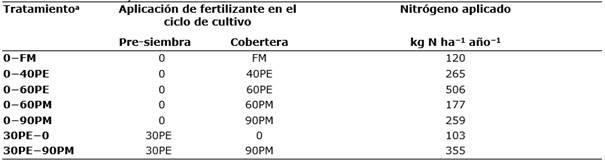
aFM, fertilización mineral; PE, purín de cerdo de engorde, los números indican las dosis teóricas de aplicación en peso fresco: 30 t ha−1 año−1 aplicado en pre-siembra, 40 y 60 t ha−1 año−1 aplicado en cobertera; PM, purín de cerdo de maternidad aplicado en cobertera, los números indican las dosis de aplicación en peso fresco: 60 y 90 t ha−1 año−1; 30PE−90PM con aporte combinado de purín de cerdo de engorde (pre-siembra) y maternidad (cobertera) a dosis de 30 y 90 t ha−1 año−1, respectivamente.
La caracterización de la comunidad de lombrices de tierra se realizó a dos niveles: (i) nivel taxonómico, que incluyó la identificación de las especies (número de especies) por la morfología externa de los especímenes adultos (Pérez y Rodríguez, 2008), (ii) nivel global, en el que se cuantificó la abundancia (individuos m-2) de juveniles y adultos y se calculó la abundancia total como la suma, considerando las lombrices enteras (cuando estaban cortadas, solo se contaba la parte anterior), y la biomasa total (g m-2) donde el peso de los especímenes presentes en las muestras y preservados en etanol (o de sus fracciones) se consideró equivalente al peso fresco.
Los resultados obtenidos se analizaron estadísticamente utilizando el programa SAS (SAS Institute 2002-2012). Se realizó un análisis de varianza (acrónimo en inglés ANOVA). Las mediciones incluidas en el nivel global se normalizaron previamente al análisis ANOVA mediante la transformación raíz cuadrada.
RESULTADOS Y DISCUSIÓN
A nivel taxonómico se identificaron dos especies de lombrices de tierra (Figuras 1 y 2). La primera especie encontrada, Koinodrilus roseus (Savigny, 1826) ha sido descrita por distintos autores, por lo que presenta distintas sinonimias: Allobophora rosea (Pop, 1948; Gerar, 1964; Alvarez, 1971; Bouché, 1972), Aporrectodea rosea (Gates, 1976; Sims y Gerar, 1985), Enterion roseum (Savigny, 1826), Eisenia rosea (Michaelsen, 1900; Gates, 1974), Allolobophora rosea (Savigny, 1826), este último mencionado en la publicación de Lainez y Jordana (1987). Pertenece al grupo de las especies endogeicas. La segunda especie encontrada, Nicodrilus trapezoides (Dugés, 1828) también presenta distintas sinonimias: Allolobophora trapezoides (Rosa, 1886), Allolobophora calaginoza trapezoides (Alvarez, 1971), Nicodrilus caliginosus meridionalis (Bauché, 1972). Pertenece al grupo de las especies anécicas.
Las especies de la familia Lumbricidae son consideradas como ingenieros del ecosistema, por las actividades de bioturbación que benefician los servicios ecosistémicos del suelo (Jones, Lawton & Shachak, 1994) y han sido utilizadas en biomonitorización de la calidad del suelo (Tischer, 2008). Las especies observadas (K. roseus y N. trapezoides) son típicas de tierras cultivadas y han sido descritas en campos de cultivos y pastizales en diversas zonas españolas (Hernández, Gutiérrez, Ramajo, Trigo & Díaz Cosín, 2003; Lainez y Jordana 1987; Pérez y Rodríguez, 2008; Pérez y Rodríguez, 2009). Las lombrices identificadas, K. roseus y N. trapezoides, son morfológica y funcionalmente distintas (Figura 1 y 2), destacando la diferencia en el tamaño del cuerpo. Estos aspectos son de relevancia porque contribuyen de manera diferenciada en la bioturbación del suelo (Pirón et al., 2017) ya que las endogeicas forman amplios sistemas de madrigueras horizontales, mezclando fragmentos de materia orgánica con la fracción mineral del suelo (son comedoras de suelo), mientras que las anécicas forman galerías verticales que facilitan el transporte de agua y nutrientes (u otras sustancias químicas) hacia capas más profundas.
Nuestros resultados muestran una baja diversidad de lombrices (2 especies identificadas) de manera similar a otros estudios en suelos agrícolas del Mediterráneo (Andriuzzi, Pulleman, Cluzeau & Pérez, 2017; Baldivieso-Freitas et al., 2018). De hecho, según Rutgers et al. (2016), en condiciones Mediterráneas semiáridas es característico encontrar 2-3 taxones y ello se explica porque las bajas precipitaciones y el bajo contenido de materia orgánica les afectan negativamente. Además, en comparación con pastizales, la diversidad disminuye con la intensificación agrícola (Curry, 2004; Rutgers et al., 2016).
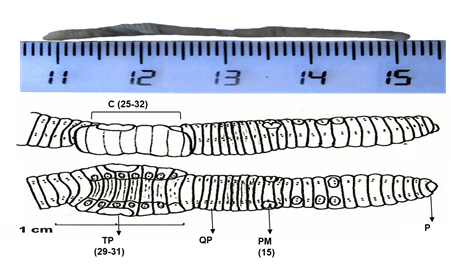
Figura 1 Características de diagnóstico (1) y generales (2) de la especie Koinodrilus roseus. (1) quetas pareadas (QP), poros masculinos (PM) en el segmento 15, pigmentación dorsal sin bandas, prostomio (P) epilóbico cerrado, clitelo (C) del 25-32, tubérculos pubertarios (TP) 29-31, primer poro dorsal en 4/5, coloración rosa en vivo (2) número de segmentos del cuerpo 120-165, longitud 25-70 mm, anchura 2-4 mm, poros nefridianos en la línea área B cerca de la línea b de quetas, poros de espermatecas en los surcos intersegmentarios 9/10 y 10/11 área D, poróforos en general si rebasar el segmento 15. Imágenes dorsales y ventrales adaptadas de Pérez y Rodríguez (2008).
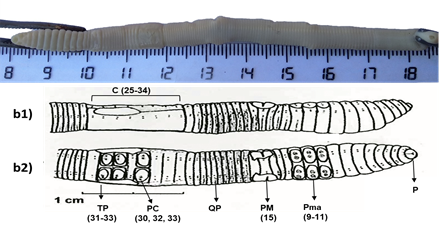
Figura 2 Características de diagnóstico (1) y generales (2) de la especie Nicodrilus trapezoides. (1) Quetas pareadas (QP), poros masculinos (PM) en el segmento 15, pigmentación dorsal sin bandas, prostomio (P) epilóbico cerrado, clitelo (C) del 25-34, tubérculos pubertarios (TP) 31-33 sin estrechamiento en 32, papilas mamelonares (PM) en los segmentos 9-11 (Pma), papilas clitelares (PC) en la quetas ab de 30, 32 y 33, primer poro dorsal en 7/8 o 8/9 o 9/10 pigmentación dorsal parda (2) número de segmentos del cuerpo 122-188, longitud 55-148 mm, anchura 3-5 mm, poróforos ½ 14-½ 16. Imágenes dorsales y ventrales adaptadas de Pérez y Rodríguez (2008).
La especie endogeica K. roseus (Figura 1) fue la dominante y representó entre el 80 y 100% de los individuos (Figura 3), independientemente de la naturaleza del fertilizante (orgánico o mineral). Por otra parte, la especie anécica N. trapezoides (Figura 2), además de minoritaria, no se registró en los tratamientos 0-40PE y 0-60PE (Figura 3). Cuando el material orgánico se entierra, las anécicas tienen desventajas (por la competencia por recursos alimenticios) frente a la endogeicas (Capowiez et al., 2009). No obstante, ello no explica la ausencia de N. trapezoides en 0−40PE y 0−60PE ya que la incorporación de residuos de cosecha es similar para todos los tratamientos. Las mayores aportaciones de N en estos tratamientos (265 y 506 kg N ha-1), tampoco justifican la desaparición de N. trapezoides ya que cuando la aportación se realiza como PM a dosis de 259 kg N ha-1 (0-90PM) o incluso combinada con PE a dosis de 355 kg N ha-1 (30PE-90PM), la especie sigue presente (Figura 3). Estos resultados indican la existencia de otros componentes diferenciales asociados al origen del purín que inciden sobre las especies y que no han sido analizados. Así, K. roseus, ha sido descrita como tolerante a distintas perturbaciones edáficas (Koblenz et al., 2015; Murchie et al., 2015) por lo que la expresión de esta tolerancia concuerda con nuestros resultados ya que esta especie está presente en todos los tratamientos (Figura 3).
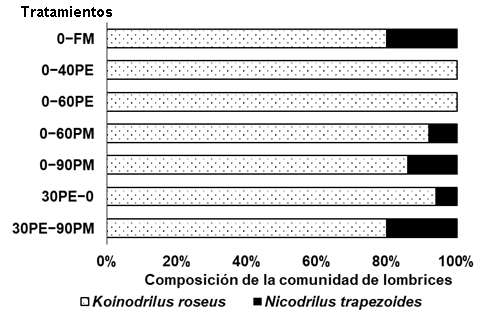
Figura 3 Composición porcentual de las dos especies de lombrices de tierra según los distintos tratamientos de fertilización: FM, fertilización mineral; PE, purín de cerdo de engorde, los números indican las dosis teóricas en peso fresco aplicadas: 30 t ha−1 año−1 aplicado en pre-siembra, 40 y 60 t ha−1 año−1 aplicado en cobertera; PM, purín de cerdo de maternidad aplicado en cobertera, los números indican las dosis en peso fresco aplicadas: 60 y 90 t ha−1 año−1; 30PE−90PM con aporte combinado de purín de cerdo de engorde (pre-siembra) y maternidad (cobertera) a dosis de 30 y 90 t ha−1 año−1, respectivamente.
La abundancia registrada (Figura 4) es la característica de zonas Mediterráneas de secano que suele oscilar entre 5−200 individuos m−2 (Rutgers et al., 2016). La abundancia (total y de las formas juveniles y adultas) así como la biomasa no variaron significativamente con la aplicación, durante 12 años, de los distintos tratamientos de fertilización (Tabla 2).
Tabla 2 Análisis de varianza de los datos normalizados (transformación raíz cuadrada) de la abundancia (juveniles, adultos e individuos totales) y de la biomasa de lombrices de tierra para los diferentes tratamientos de fertilización.
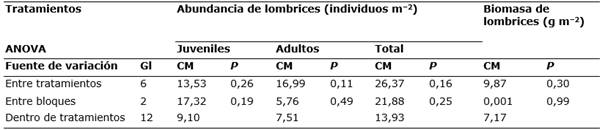
a Gl= grados de libertad, CM= cuadrados medios, P= valor de probabilidad.
En general, los tratamientos asociados a la fertilización orgánica previa presentaron, respecto a la fertilización mineral (0−FM), valores de abundancia total entre un 21 a un 310% superiores (rango de 48 a 197 individuos m−2), y también superiores (entre 68 a 494%) en biomasa (rango de 9,0 a 53,5 g m−2) (Tabla 3). Estas diferencias se observaron tanto en las formas adultas como en las juveniles (Tabla 3).
Tabla 3 Valores medios y error estándar de la abundancia (juveniles, adultos e individuos totales) y biomasa de lombrices de tierraa para los diferentes tratamientos de fertilización.
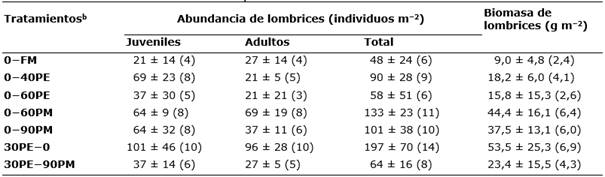
aLos números entre paréntesis corresponden a los datos normalizados mediante la transformación de la raíz cuadrada.
bFM: fertilización mineral; PE: purín de cerdo de engorde, los números indican las dosis, en peso fresco,teóricas: 30 t ha-1 año−1 aplicado en pre-siembra, 40 y 60 t ha-1 año−1 aplicadas en cobertera; PM: purín de cerdo de maternidad, los números indican las dosis, en peso fresco, de aplicación en cobertera: 60 y 90 t ha-1 año−1; 30PE−90PM con aporte de purín de cerdo de engorde (pre-siembra) y maternidad (cobertera) a dosis de 30 y 90 t ha-1 año−1, respectivamente.
La tendencia mostrada en los parámetros evaluados en la fertilización con purines respecto a la mineral se explicaría porque las aplicaciones a largo plazo de fertilizantes orgánicos incrementan las fuentes de alimentos (nutrientes y MO), beneficiando directamente e indirectamente (v.g. mejora de la estructura y porosidad, favoreciendo su hábitat) a la comunidad de lombrices de tierra (Biau, Santiveri, Mijangos & Lloveras, 2012; Baldivieso-Freitas et al., 2018) lo que repercute en un incremento de la abundancia y la biomasa (Hansen & Engelstad, 1999; Baldivieso-Freitas et al., 2018).
El estudio confirma los efectos beneficiosos del purín de cerdo sobre la comunidad de lombrices de tierra a dosis de N próximas al umbral máximo (170 kg N año-1 ha-1 de origen orgánico) establecido para zonas susceptibles de contaminación de las aguas subsuperficiales por nitratos (Unión Europea, 1991). El estudio también indica que el uso de dosis más elevadas de purín tiende a disminuir significativamente la abundancia y biomasa de lombrices (P= 0,02 y P= 0,03, respectivamente) pero siempre con valores similares o por encima de la fertilización mineral (Figura 4). Varios autores (Curry, 1976; Hansen & Engelstad, 1999; Edwards, 2007; Koblenz et al., 2015) afirman que los efectos negativos de la sobrefertilización con purines sobre la comunidad de lombrices de tierra (menor abundancia y biomasa) están relacionados con el efecto tóxico del N-amoniacal y otras sales, pero que el estrés se produce generalmente a corto plazo (inmediato a la aplicación) y es transitorio, por lo que la población podría recuperarse si las aplicaciones no fueran frecuentes. Sin embargo, esto no se evidencia en este ensayo puesto que se ha evaluado la población de lombrices tras un periodo superior a los 12 meses desde la última aplicación de purín, y el N-amoniacal en este tipo de suelo se transforma rápidamente en N-nítrico (Sánchez y González, 2005). Otros autores (Ma, Brussaard & de Ridder, 1990) asocian la sobrefertilización con la disminución del pH y consecuente efecto negativo en las lombrices. Pero en estudios previos, en este mismo experimento (Bosch-Serra, Padró, Boixadera-Bosch, Orobitg & Yagüe, 2014), no se observaron cambios en el pH (permanecía entre 8,3−8,4) a dosis crecientes de purín porcino debido al alto poder tampón de los suelos ricos en carbonato de calcio.
Por otro lado, una sobrefertilización con purines también implica riesgos ecotoxicológicos (De la Torre et al., 2000). Los metales pesados podrían acumularse en el suelo tras aplicaciones a largo plazo de purín y aunque no se alcanzasen niveles fitotóxicos (Berenguer, Cela, Santiveri, Boixadera & Lloveras, 2008), las lombrices de tierra podrían estar en riesgo (Becquer et al., 2005) debido a su hábito alimenticio (ingestión de suelo y MO). Esta situación podría darse incluso cuando los residuos orgánicos cumpliesen con los requisitos para uso agrícola (Renaud et al., 2017). Estos potenciales riesgos ecotoxicológicos requerirían de mayor experimentación.
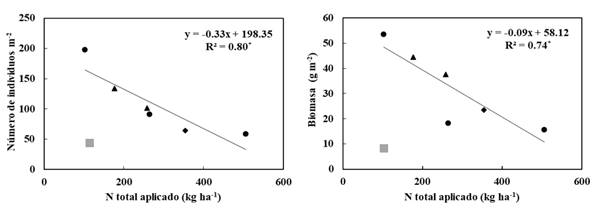
Figura 4 Relación entre la abundancia (individuos totales) y biomasa de lombrices de tierra y la dosis de N aplicada (* P<0,05) cuando se fertiliza con purines. Se señalan las aplicaciones de N como fertilizante mineral (cuadrado en gris), como purín de cerdo de engorde (circulo en negro), como purín de madres reproductoras (triángulo en negro) y la aplicación combinada de purín de engorde y madres reproductoras (rombo en negro).
CONCLUSIONES
En una situación de barbecho, tras doce años de fertilización con purín porcino y en relación a la fertilización mineral, la población de lombrices de tierra a nivel global (abundancia y biomasa) no experimenta cambios significativos. La abundancia total oscila entre 48 y 197 individuos m−2 y la biomasa entre 9 y 54 g m−2. Se identificaron dos especies: Koinodrilus roseus que fue la dominante y Nicodrilus trapezoides. La especie Nicodrilus trapezoides no se detecta en los muestreos hasta 20 cm en condiciones de sobrefertilización histórica con PE.
La fertilización nitrogenada con dosis ajustadas de purín porcino, cualquiera sea su origen (PE o PM), presenta el mayor número de lombrices del suelo y diversidad, por lo que la dosis de N recomendada por la Unión Europea (170 kg N ha−1 año−1 de fuentes orgánicas) para zonas clasificadas como vulnerables a la contaminación de las aguas subterráneas por nitratos, sería también la adecuada en relación a la prevención de riesgos ambientales ligados a la pérdida de diversidad edáfica. La ausencia de N. trapezoides en los primeros 20 cm puede considerarse un buen indicador de aportes excesivos de purines.
Serían necesarias futuras investigaciones para determinar los componentes del purín de cerdos de engorde que impactan negativamente sobre las lombrices de tierra al incrementar las dosis aplicadas.