











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
El rendimiento de Hordeum vulgare (cebada) disminuye por pérdida de fertilidad del suelo (Contreras-López et al. 2008); consecuencia en parte por prácticas culturales inadecuadas como riego superficial y sobreexplotación, así como el uso excesivo de fertilizante nitrogenado (FN), como NH4NO3 (nitrato de amonio) empleado en su cultivo, lo que provoca pérdida de la materia orgánica y contaminación ambiental (Palafox-Caballero et al. 2005). Una opción para regular y optimizar el uso de FN es la inoculación de las semillas de cebada con bacterias promotoras del crecimiento vegetal endófitas (BPCVE), que al colonizar el interior de sus raíces y el sistema de conducción vegetal, estimulan la formación de raíces primarias y secundarias adecuada para su crecimiento, y facilitan la absorción del FN (James 2000).
Las BPCVE tienen mayor disponibilidad de nutrientes en el interior de los tejidos vegetales y mínima competencia con otros géneros bacterianos relacionados, tal como sucede en la rizósfera y suelo (James 2000; Muthukumarasamy et al. 2002). Tal es el caso de los géneros de BPCVE Burkholderia cepacia y Gluconacetobacter diazotrophicus que invaden el sistema de conducción y en el interior del sistema radical, en donde los carbohidratos y ácidos orgánicos, metabolitos generados por la fotosíntesis les sirven como fuente de alimento; cuando colonizan y circulan en el xilema de una amplia variedad de plantas domésticas y silvestres (Sevilla et al. 2001;Youssef et al. 2004; Muñoz-Rojas et al. 2005). Las BPCVE pueden transformar esos metabolitos de la fotosíntesis en sustancias promotoras del crecimiento vegetal (SPCV) (Triplett 2000; Riggs et al. 2001), con las que ejercen efectos positivos en el crecimiento de gramíneas como la cebada.
Las BPCVE tienen una quimiotaxis positiva por los exudados orgánicos del sistema radical de la cebada, que para su conversión en SPCV en la proliferación de pelos radicales, los que en consecuencia aumentan el área de exploración y absorción del FN (Rojas et al. 2009), principalmente cuando su concentración se reduce. Como lo observado en otras Poaceas, al respecto Plana et al. (1999), reportaron la respuesta positiva de Triticum aestivum a B. cepacia y Azospirillum brasilense en función de su peso seco total (PST) con aumento del 20%, en relación con el mismo PST de T. aestivum con la dosis del FN recomendada y sin tratar con esas BPCVE. Mientras que García-González et al. (2005), reportaron un incremento entre el 15 al 25% del peso fresco total (PFT) y PST de T. aestivum inoculado con Azospirillum spp. y Azotobacter beijerinckii a dosis 50% del FN respecto a la misma variable en T. aestivum usado como CR a la dosis del FN sugerido en la región, sin las BPCVE. Igualmente, Mora y Toro (2007) evaluaron el efecto positivo de Burkholderia sp. endófita en Zea mays mediante la síntesis de SPCV que indujeron un incremento en la longitud radical y biomasa entre el 28 y 52% respectivamente, en comparación con Z. mays empleado como CR a la dosis de FN recomendada en la región. A pesar de lo anterior, en la literatura no se reporta el uso de B. cepacia ni G. diazotrophicus, géneros de BPCVE en cebada a dosis 50% del FN.
El objetivo de este trabajo fue analizar la respuesta de cebada a la inoculación con Burkholderia cepacia y Gluconacetobacer diazotrophicus.
MATERIALES Y MÉTODOS
Se utilizó un suelo ubicado a los 19º 39' 27'' de latitud norte y 100º 19' 59'' de longitud oeste, con una altitud de 1.820 msnm clima templado, temperatura media anual de 17,3ºC, precipitación anual de 796 mm, granizadas promedio 3-4/año, heladas 4-8/año e insolación 227:63 h:min; en un terreno agrícola denominado "La cajita'' de la Tenencia Zapata del municipio de Morelia, Mich., sobre el km 5 de la carretera Morelia- Pátzcuaro, México.
El suelo fue solarizado para minimizar el problema de plagas y enfermedades (Sánchez-Yáñez 2007). El área cuenta con un historial agrícola de 20 años con sistema de cultivo intensivo cereal-cereal: maíz-trigo y maíz-cebada. El suelo fue tipificado como lateritico sódico según la clasificación de la FAO (2007): de textura arcillosa, con bajo contenido de materia orgánica (15 g kg-1), y de N orgánico (39 kg ha-1), pH 6,7 ligeramente ácido (Whitmore 2000).
El género y especie de B. cepacia se aisló de las raíces de Z. mays sp. mexicana (teocintle) en un medio de cultivo conocido como agar Pseudomonas cepacia ácido azelaico y triptamina (APCAT) (Tran Van et al. 2000). Este aislado se identificó mediante su perfil bioquímico con el sistema API-50CH (BioMerieux, Francia).
Según se describe en la literatura, B. cepacia endófita de Z. mays sp. mexicana, se comparó con las siguientes cepas de referencia: B. cepacia ATCC 2546 y B. vietnamiensis TVV75. En relación al aislado de G. diazotrophicus se recuperó de Saccharum officinarum en caldo y Agar Lisina Glucosa Indol (ALGI) incubado a 32ºC por 24 h (Youssef et al. 2004; Dibut et al. 2005); este aislado se identificó a nivel de especie con el sistema API20NE (Salazar et al. 2008). Para demostrar que B. cepacia y G. diazotrophicus tenían la capacidad de sintetizar SPCV, en específico algún tipo de giberelina, cada uno se sembró en APCAT y AGLI. Para ello se suprimió el agar y en medio líquido se cultivó cada género de BPCVE en matraces de 250 mL con 100 mL APCT y AGLI. Los matraces se agitaron a 250 rpm con un mínimo de 24 h y un máximo de 48 h, temperatura 30°C en función del crecimiento suficiente para iniciar la inoculación de la semilla de cebada. Posteriormente cada matraz se centrifugó a 3.000 rpm/15 min; luego se eliminó el paquete celular y el sobrenadante se filtró con una membrana milipore 0,2 µ estéril (Gelman, Co); para obtener el filtrado estéril libre de células de B. cepacia y G. diazotrophicus: que contenían las SPCV relacionadas con algún de tipo giberelina.
Como referencia positiva para la germinación de las semillas se aplicaron 25 ppm de giberelina (Sigma, Co) pura disuelta en agua estéril y como control absoluto (CA); agua estéril. Con base en los siguientes tratamientos: a) los filtrados estériles libres de células de B. cepacia y G. diazotrophicus; b) giberelina pura; c) agua estéril. Se emplearon 100 semillas de H. vulgare, P. vulgaris y Z. mays respectivamente; colocadas en caja de Petri sobre algodón humedecido esterilizado, mantenidas a temperatura entre 25-30°C/9 días. Para este ensayo se midieron las variables respuesta: porcentaje y días de germinación (Singh et al. 2003; García-González et al. 2005; Tsavkelova et al. 2006); Burkholderia cepacia y Gluconacetobacter diazotrophicus fueron inoculados en la semilla de cebada, para lo que por cada 200 semillas se les aplicó 1,0 mL de B. cepacia o G. diazotrophicus a una densidad de 600x106 UFC mL-1 calculado con base en una cuenta viable en placa en APCAT y ALGI respectivamente. Cuando se realizó la doble inoculación con B. cepacia y G. diazotrophicus por cada 200 semillas de cebada se usó 1,0 mL-1 (0,5 mL de cada una) en una proporción 1:1 (Bashan y Levanony 1998; García-González et al. 2005).
En la Tabla 1 se muestran los tratamientos utilizados para medir el efecto de las BPCVE en H. vulgare var. Armida a la dosis 50% del FN como NH4NO3 5 g L-1. En Michoacán, México para la cebada se recomienda una dosis de FN equivalente a 10 g L-1 de NH4NO3 (Cortés y Ortiz 2008; Villaseñor et al. 2014). Este ensayo se realizó en invernadero, ahí el suelo se colocó en el sistema hidropónico de jarras de Leonard (García-González et al. 2005); en el reservorio de cada jarra de Leonard se agregó la SM con la siguiente composición química (g L-1): NH4NO3 10,0; K2HPO4 2,5; KH2PO4 2,0; MgSO4 1,0; NaCl 0,1; CaCl2 0,1; FeSO4 trazas, ajustado a pH 6,5 y 10,0 mL de solución de oligoelementos con la siguiente composición (g L-1): H3BO3 2,86, ZnSO47H2O 0,22: MnCl27H2O 1,81; K2MnO4 0,09; ajustado a pH 6,8; se usaron 500 mL/15 días/jarra de Leonard. En el suelo de cada jarra de Leonard, la humedad se mantuvo al 80% de capacidad de campo, con agua potable por 14 semanas (Sánchez-Yáñez 2007).
Los tratamientos fueron dispuestos bajo un diseño experimental unifactorial de seis tratamientos y seis repeticiones en cada uno: a) cebada sin inocular, sin FN (control absoluto, CA); b) cebada sin inocular con el 50% de FN (control relativo 1, CR1); c) cebada sin inocular con el 100% de FN (control relativo 2, CR2), d) cebada inoculada con B. cepacia y el 50% de FN; e) cebada inoculada con G. diazotrophicus y 50% del FN y f) cebada inoculada con la mezcla de B. cepacia y G. diazotrophicus y el 50% del FN (García-González et al. 2005).
En la Tabla 1 se muestra el diseño experimental empleado para evaluar la respuesta de la cebada a la inoculación con B. cepacia y G. diazotrophicus a nivel de plántula y de floración, 32 y 62 días después de la siembra. Las variables respuesta evaluadas fueron: altura de la planta (AP) y longitud de la raíz (LR), ambas medidas tomadas con una regla calibrada. Para la AP se consideró desde la base del cuello hasta el ápice máximo de crecimiento; para la LR se tomó desde el cuello de la planta a la longitud máxima de la raíz principal. También fueron registrados los valores para biomasa: peso fresco total (PFT), para ello se tomó una planta recién colectada, se seleccionó la parte aérea que se pesó en una balanza electrónica, y para el peso seco total (PST) la planta se secó en un horno a 75°C/30 h hasta registrar un peso constante.
Los datos experimentales se analizaron por ANOVA y Tukey con un nivel de significancia de P>0,05 para establecer las diferencias en los resultados obtenidos, cuando la cebada fue inoculada con B. cepacia y/o G. diazotrophicus a la dosis 50% del FN, en comparación con la cebada sin inocular con el FN al 100% (García et al. 2001).
RESULTADOS Y DISCUSIÓN
Efecto de Burkholderia cepacia y Gluconacetobacter diazotrophicus fue observado en la cebada con el FN al 50% (Figura 1), en las fases de plántula y floración; donde se verificó que B. cepacia registró un valor de 43,75 cm de la AP, sin diferencia estadística con los 41,0 cm de la cebada con la dosis 100% de FN o CR; pero sí con los 31,12 cm de AP de la cebada irrigada con agua o CA. Esto sugiere que el efecto positivo de B. cepacia en el interior del tejido de raíz de la cebada, fue el resultado de la transformación de algunos de los compuestos orgánicos derivados de la fotosíntesis en SPCV para su sano crecimiento (García et al. 2001).
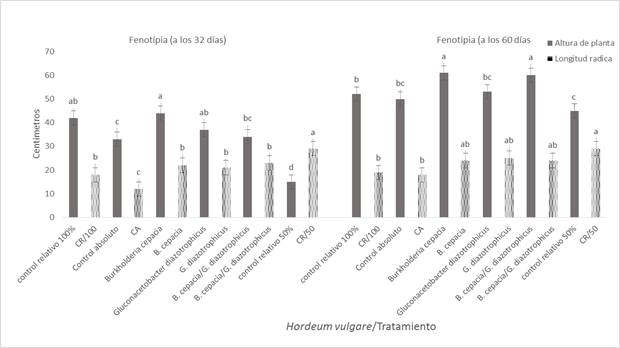
Figura 1 Efecto de la inoculación de Burkholderia cepacia y Gluconacetobacter diazotrophicus en Hordeum vulgare var. Armida en la fenología a plántula y floración.
La respuesta positiva de la cebada a B. cepacia y G. diazotrophicus, en relación a LR, registraron un valor de 23,5 cm, sin diferencia estadística con los 18,5 cm con la LR de la cebada alimentada con la dosis 100% del FN o CR. Sin embargo, ambos valores fueron estadísticamente diferentes e inferiores a los 12,0 cm de la cebada irrigada solo con agua o CA. Este resultado indica que los dos géneros de BPCVE mediante SPCV, indujeron una mayor proliferación de raíces secundarias que incrementó su capacidad de absorción radical del FN, a pesar de reducir la dosis al 50%. Este fue un resultado positivo, análogo a lo reportado en Z. mays inoculado con la endófita Burkholderia sp. con la dosis del FN inferior a lo recomendado para esa gramínea (Riggs et al. 2001; Mora y Toro 2007).
Se observó un efecto positivo de B. cepacia en la cebada a floración con una AP de 61,0 cm, así como con los 58,62 cm de la cebada tratada con B. cepacia y G. diazotrophicus, ambos valores fueron estadísticamente diferentes y superiores en comparación con los 52,0 cm registrados en el tratamiento CR y con los 49,0 cm del CA. Estos resultados sugieren que en el xilema de la planta, B. cepacia como G. diazotrophicus actuaron simultáneamente en la generación de SPCV, y con ello para una eficaz absorción radical del FN a pesar de reducirlo al 50%. También se observó un efecto benéfico de G. diazotrophicus en la LR de la cebada, con 25,33 cm; al igual que con los 24,56 cm de la planta tratada con B. cepacia, ambos valores fueron estadísticamente diferentes y superiores respecto a los 19,3 cm de la LR obtenida en el CR y los 18,5 cm del tratamiento solo con agua o CA. Estos resultados sugieren que los dos géneros de BPCVE, al colonizar y proliferar en el tejido interno de la raíz de la cebada, convirtieron algunos de los aminoácidos derivados de la fotosíntesis en SPCV lo que optimizó el FN a pesar de reducirlo al 50% (Chanway et al. 2000; Bent et al. 2001; Muthukarasamy et al. 2002; Rojas et al. 2009).
Efecto positivo de Burkholderia cepacia y Gluconacetobacter diazotrophicus en Hordeum vulgare var. Armida fue observado en la variable analizada PFT, en especial con B. cepacia, en la que se obtuvo 7,78 g, valor estadísticamente diferente y superior a los 3,52 g obtenido con la dosis del FN al 100% o CR y los 1,87 g con el tratamiento irrigado solo con agua o CA. Esto puede indicar que B. cepacia en el interior de la raíz de la cebada, convirtió algunos compuestos orgánicos derivado de la fotosíntesis en SPCV, causando un incremento de la absorción y optimización de la dosis del FN al 50%, sin afectar su crecimiento (García et al. 2001). Además, B. cepacia en la cebada registró un PST de 0,85 g valor sin diferencia estadística en comparación con los 0,54 g del tratamiento con el 100% del FN o CR; estos valores fueron estadísticamente distintos a los 0,23 g del CA (Figura 2).
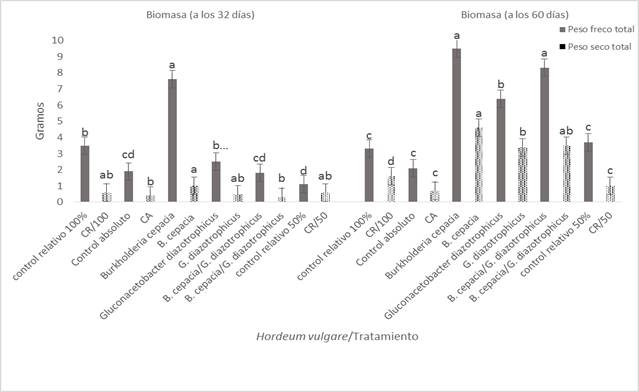
Figura 2 Efecto de la inoculación de Burkholderia cepacia y Gluconacetobacter diazotrophicus en Hordeum vulgare var. Armida en su biomasa a plántula y floración.
B. cepacia registró 9,62 g de PFT y G. diazotrophicus 8,22 g, ambos valores sin diferencia estadística en comparación con el PFT de la cebada del CR, lo que sugiere que en el xilema de esta planta las BPCVE mediante la generación de SPCV mejoraron y optimizaron la dosis del FN al 50% (Mora y Toro 2007). La inoculación de BPCVE en Z. mays indujo un incremento en la formación de raíces secundarias, con un aumento en la capacidad de absorción radical del FN al 50%, sin que el resultado haya sido un crecimiento anormal del Z. mays, en comparación con el Z. mays tratado con el FN al 100% sin inocular con las BPCVE (Riggs et al. 2001; Mora y Toro 2007). Estos valores de PFT (9,62 y 8,22 g) fueron estadísticamente diferentes y superiores a los 3,22 g de PFT obtenido con la dosis del FN al 100% o CR y los 2,0 g del tratamiento irrigado con agua o CA.
Cuando B. cepacia se inoculó en la cebada se registró 4,48 g de PST, mientras que con la mezcla de B. cepacia y G. diazotrophicus se obtuvo 3,29 g para la misma variable. Ambos valores fueron estadísticamente diferentes y superiores a los 1,49 g de PST obtenido en el CR y los 0,66 g del CA. Este hecho apoya que en el xilema de la cebada, tanto B. cepacia como G. diazotrophicus sintetizaron SPCV, que aumentaron su capacidad de absorción radical del FN al 50% para su optimización y permitir su sano crecimiento (Chanway et al. 2000; Muthukarasamy et al. 2002). Dado que B.cepacia como G. diazotrophicus endófitos de plantas, tienen la capacidad genética de convertir metabolitos de la fotosíntesis en SPCV, con una acción similar a la de ciertas auxinas y giberelinas (Bent et al. 2001), específicamente cuando géneros y especies de BPCVE se asocian con las Poaceas del tipo S. officinarum y lo novedoso e interesante también con H. vulgare (Rojas et al. 2009) de acuerdo a los resultados obtenidos en este investigación.
CONCLUSIONES
Las BPCVE como B. cepacia causan un efecto positivo en el crecimiento de la cebada, no obstante disminuir la dosis del FN al 50%, en especial en comparación a lo que se registra cuando se inocula con G. diazotrophicus.
El hecho de que ambos géneros de BPCVE colonicen el xilema de la planta, sugiere que la respuesta positiva de la cebada es relativamente posible cuando se inocula con B. cepacia, a pesar de reducir la dosis del FN al 50%, inferior a lo recomendado de acuerdo a la variedad de cebada y tipo de suelo, sin afectar negativamente su crecimiento, lo que aunado a prácticas culturales de conservación agrícola evitan la perdida de la fertilidad del suelo.

