








Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkInvestigación Agraria
versión On-line ISSN 2305-0683
Investig. Agrar. vol.18 no.1 San Lorenzo jun. 2016
https://doi.org/10.18004/investig.agrar.2016.junio.30-36
ARTÍCULO CIENTÍFICO
Parámetros biológicos de Tetranychus gloveri Banks (Acari: Tetranychidae) sobre hojas de Cocos nucifera L.
Biological parameters of Tetranychus gloveri Banks, 1900 (Acari: Tetranychidae) on Cocos nucifera L. leaves
Hernán Zurita1, María A. Martínez2, Carlos Vásquez1,2*
1 Facultad de Ciencias Agropecuarias, Universidad Técnica de Ambato, Campus Querochaca, Cantón Cevallos, Tungurahua, Código postal: 18-01-334. Ecuador.
2 Universidad Centroccidental Lisandro Alvarado. Decanato de Agronomía. Departamento de Ciencias Biológicas.
* Autor para correspondencia (ca.vasquez@uta.edu.ec)
Recibido: 16/03/2016; Aceptado: 27/04/2016.
RESUMEN
Tetranychus gloveri Banks ha sido registrado atacando 107 especies de plantas y generalmente es considerada una plaga severa. Se realizaron estudios en laboratorio para determinar el ciclo de vida, la fecundidad, longevidad y parámetros de la tabla de vida de Tetranychus gloveri Banks sobre hojas de Cocos nucifera L. Los estudios biológicos se realizaron en hojas aisladas de coco var. Enano Amarillo Malayo (28 ± 3°C, 70 ± 10% HR). El tiempo total de desarrollo duró 12,0 ± 0,95 días y los tiempos promedio de preoviposición, oviposición y postoviposición fueron 3,0 ± 1,11; 9,33 ± 3,57 y 1,22 ± 0,67 días, respectivamente. La fecundidad promedio fue 29,8 huevos hembra-1 y la tasa de oviposición 2,9 ± huevos hembra-1 día-1, mientras que la longevidad promedio de las hembras de T. gloveri fue 13,1 ± días. Para los parámetros de la tabla de vida se obtuvieron valores para la tasa intrínseca de crecimiento (rm) 0,100 individuos hembra-1 día-1, el tiempo generacional (T) fue de 19,0 días, la tasa neta de reproducción (R0) fue de 6,73 y la tasa finita de crecimiento natural (λ) fue de 1,105 individuos hembra-1. Los resultados de los parámetros de incremento poblacional obtenidos en el presente estudio indican que T. gloveri no expresó un alto potencial biótico al ser criado sobre hojas deC. nucifera.
Palabras clave: Tetranychus gloveri, Cocos nucifera, ácaros fitófagos, biología.
ABSTRACT
Tetranychus gloveri Banks has been reported feeding on 107 plant species and it is generally considered a severe pest. Laboratory studies were carried out to determine life cycle, fecundity, longevity and life table parameters of Tetranychus gloveri Banks on Cocos nucifera L. leaves. Biological studies were conducted on detached coconut leaves var. Malayan Yellow Dwarf (28 ± 3°C, 70 ± 10% RH). Total developmental time for T. gloveri was 12.0 ± 0.95 days and the pre-oviposition, oviposition and post-oviposition mean time were 3,0 ± 1,11; 9,33 ± 3,57 and 1,22 ± 0,67 days, respectively. Mean fecundity was 29.8 eggs female-1 and oviposition rate was 2.9 eggs female-1 day-1 while mean longevity of T. gloveri females was 13.1 days. Life table parameters showed values for intrinsic rate of natural increase (rm) 0.100 individuals/female/day, generation time (T) 19.0 days, net reproduction rate (R0) 6.73, the finite natural increase rate (λ) 1,105 individuals female-1. Population increase parameters indicated that T. gloveri did not show a high biotic potential when reared on C. nucifera leaves.
Key words: Tetranychus gloveri, Cocos nucifera, phytophagous mites, biology.
INTRODUCCIÓN
Los ácaros fitófagos constituyen un serio problema para la producción de hortalizas y de especies forestales a nivel mundial (Gerson y Weintraub 2012), principalmente aquellas especies dentro de la familia Tetranychidae, las cuales presentan amplia distribución geográfica (Bolland et al. 1998) y alto potencial biológico. El potencial biológico depende de la planta hospedera (la variedad, estado nutricional y edad de la planta), condiciones ambientales (principalmente temperatura y humedad) y de los aspectos intrínsecos de cada especie de ácaro (Gutierrez y Helle 1985; Tomczyk y Kropczynska 1985; Wrensch 1985; Adango et al. 2006; Vásquez et al. 2007).
Entre los factores intrínsecos que influencian los parámetros de tabla de vida de estos ácaros incluyen la raza y el nivel de endogamia, densidad de la colonia, edad de las hembras y de la población, y fertilidad de las hembras (Wrensch 1985). Todos estos factores influyen sobre la expresión del daño, el cual se inicia con la introducción del estilete en el sitio de alimentación y consecuente destrucción de las células del mesófilo, afectando la transpiración y la fotosíntesis y el crecimiento de la planta y sus frutos (Avery y Brigss 1968, Tanigoshi y Davis 1978; Sances et al. 1979a, b; De Angelis et al. 1983; Gallardo et al. 2005). Esto permite suponer la complejidad de las relaciones entre las diferentes especies de ácaros fitófagos y el nivel de daño que pueden causar sobre la misma planta hospedera en diferentes regiones del mundo (Monjarás-Barrera et al. 2015).
Tetranychus gloveri Banks ha sido registrado atacando 107 especies de plantas distribuidas en diferentes familias botánicas siendo reportada en la región Neotropical, ha sido registrado en Brasil, Colombia, Costa Rica, Cuba Guadalupe, Honduras, Martinica, Panamá, Paraguay, Puerto Rico, Surinam, Trinidad y Tobago y Venezuela (Migeon y Dorkeld 2015). Dentro de la familia Arecaceae, este ácaro ha sido registrado en Chamaedorea sp., Livistona sp., Phoenix sp., Roystonea regia (Kunth) O.F. Cook y Cocos nucifera L. (Flechtmann et al. 1999). A pesar de la amplia distribución y número de especies de plantas atacadas por T. gloveri, hasta el presente no existen estudios relacionados con su biología, por lo que en el presente estudio se evaluaron algunos aspectos sobre ciclo biológico de T. gloveri sobre hojas de coco.
MATERIALES Y MÉTODOS
Colecta y mantenimiento del ácaro
Los ácaros fueron colectados de plantas de coco variedad Enano Amarillo Malayo del sector Los Rastrojos, Municipio Palavecino, estado Lara, Venezuela. Las hojas fueron colocadas en bolsas plásticas de cierre hermético, internamente recubiertas con papel absorbente y llevadas en cavas refrigeradas al laboratorio de Investigación de Zoología Agrícola, Universidad Centroccidental “Lisandro Alvarado”. En el laboratorio, las hojas fueron examinadas bajo el aumento del microscopio estereoscópico para seleccionar los ácaros tetraníquidos, los cuales fueron montados en láminas microscópicas. La determinación del género se ha realizado utilizando la clave taxonómica de Gutierrez (1985), mientras que la especie fue determinada por comparación con la morfología del aedeago (Ochoa et al. 1994).
Previo al inicio del ensayo, se prepararon cinco unidades de cría para la obtención de individuos de edad homogénea, siguiendo la metodología de Helle y Overmeer (1985). Cada unidad de cría consistió en una cápsula de Petri (9 cm de diámetro) que contenía una almohadilla de poliuretano de 1 cm de espesor y humedecida con agua destilada. Sobre cada unidad de cría fueron colocados dos discos de hoja de coco (2 cm de diámetro) variedad Enano Amarillo Malayo con el envés hacia arriba, sobre las cuales fueron colocados cinco hembras y dos machos para promover la cópula y asegurar la producción de huevos. Después de 24 h las hembras y machos fueron descartados y se registró el número de huevos. Los huevos obtenidos fueron dejados sobre las unidades de cría hasta la emergencia de los adultos, los cuales fueron observados en el estudio de ciclo biológico.
Ciclo de vida de T. gloveri
El ciclo biológico fue estudiado bajo condiciones de laboratorio (28 ± 3°C, 70 ± 10% HR) en unidades de cría similares a las descritas arriba. El estudio fue iniciado con 20 unidades de cría, cada una de ellas conteniendo dos discos de hoja colocados con la superficie abaxial hacia arriba; que fueron rodeados de una banda de algodón humedecida para evitar el escape de los ácaros y mantener la turgencia de la hoja. Sobre cada disco de hoja fueron colocados una hembra y un macho (de aproximadamente 2 días de edad) provenientes de la cría del laboratorio.
Las unidades de cría fueron observadas cada 12 h usando un microscopio estereoscopio para determinar el momento de la oviposición, luego fueron eliminados hembras y machos de cada unidad. La determinación del tiempo de incubación y duración de las diferentes fases (larva, protocrisálida, protoninfa, deutocrisálida, deutoninfa y teliocrisálida) fueron observados a intervalos de 12 h.
Preoviposición, oviposición, postoviposición, fecundidad y longevidad de T. gloveri
Los períodos de preoviposición, oviposición y postoviposición fueron estudiados en un grupo de 20 hembras. Cada hembra fue colocada individualmente en una unidad de cría previamente identificada y observada cada 24 h para determinar período de preoviposición (tiempo transcurrido entre la emergencia de la hembra hasta el momento de la primera oviposición), así como el tiempo de oviposición (tiempo desde la primera oviposición hasta el último huevo) y el período de postoviposición (tiempo desde que coloca el último huevo hasta la muerte de la hembra). Adicionalmente se contabilizó el número de huevos por hembra (fecundidad) y el tiempo desde la emergencia de la hembra hasta la muerte (longevidad). Los datos fueron expresados en días promedio para cada uno de los períodos. La longevidad se determinó desde la emergencia de las hembras adultas hasta la muerte.
Tabla de vida de T. gloveri
Los parámetros de la tabla de vida fueron calculados a partir de los datos de sobrevivencia de las hembras y proporción de descendencia por cada hembra, obtenidos en el laboratorio según Birch (1948). Se estimó el tiempo generacional (T), reproducción neta (Ro), la tasa intrínseca de crecimiento (rm) y la tasa de incremento finito mediante el uso de las fórmulas siguientes:
a. Tasa Intrínseca de Crecimiento (rm): se define como la capacidad de multiplicación de una población en el lapso de una generación.
Donde: x: = edad de los individuos en días. lx = la proporción de individuos vivos a la edad 'x'. mx = número de descendencia hembra producida por cada hembra en el intervalo de edad 'x'.
b. Tiempo Generacional (T): representa el tiempo promedio entre dos generaciones sucesivas.
c. Tasa Neta de Reproducción (R0): conocido usualmente como tiempo de reemplazo. Este refleja el número promedio de progenie hembra que es capaz de producir cada hembra de la población durante su vida.
d. Tasa Finita de Crecimiento Natural (l): se interpreta como el número de individuos que se agrega a la población por individuo y por unidad de tiempo.
Análisis estadístico
Los ensayos de duración del ciclo de vida, fecundidad y longevidad de T. gloveri fueron conducidos en un diseño completamente al azar y expresados como valores promedios.
RESULTADOS Y DISCUSION
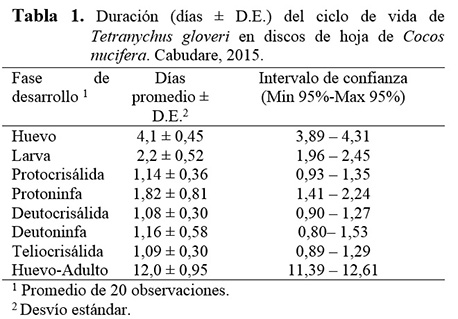
Duración del ciclo biológico de T. gloveri
El tiempo promedio de desarrollo de T. gloveri desde la fase de huevo hasta la emergencia de la fase adulta fue de 12,0 ± 0,95 días, la fase de huevo duró un promedio de 4,1 ± 0,45 días, el estadio larval tuvo una duración promedio de 2,2 ± 0,52 días. Las fases intermedias tuvieron una duración en protocrisálida de 1,14 ± 0,363, protoninfa de 1,82 ± 0,810, deutocrisálida de 1,08 ± 0,289, deutoninfa de 1,16 ± 0,577, teliocrisalida de 1,09 ± 0,302 días respectivamente (Tabla 1). Hasta el presente, no existen estudios disponibles sobre ciclo biológico de T. gloveri, por lo que los resultados fueron comparados con otras especies del género Tetranychus. En tal sentido, Stein y Daólio (2012) reportaron que Tetranychus mexicanus (McGregor) presentó un ciclo de vida de 13,6 días sobre discos de hoja de Bactris gasipaes Kunth (Arecaceae) a 25°C, 60% de humedad relativa y 14 h de fotoperíodo. Otros trabajos han señalado que el tiempo de duración del ciclo biológico de T. urticae varió desde 15,7 días sobre plantas de gerbera a 25°C (Silva et al. 2009) hasta 7-8 días sobre hojas de Lablab purpureus L. a 26,03°C (Fazlul-Hoque et al. 2008). En general, el tiempo de desarrollo de los tetraníquidos es afectado por factores relacionados con la temperatura, humedad, depredación, competencia inter-específica, plaguicidas y características de la planta hospedante, así como por factores intrínsecos de la especie particular de ácaro (Vrie et al. 1972, Wrensch 1985).
Tiempo de preoviposición, oviposición, postoviposición, fecundidad y longevidad de T. gloveri
La duración promedio del período de preoviposición fue de 3,0 días, la duración de los periodos de oviposición y post-oviposición fueron de 9,33 y 1,22 días, respectivamente, el tiempo promedio de vida de las hembras de T. gloveri fue de 13,1 días (Tabla 2).
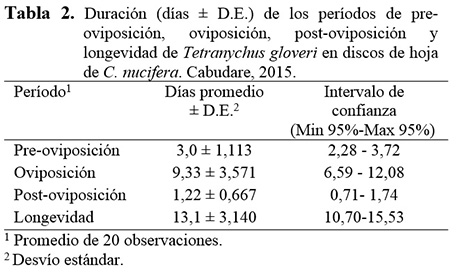
Con relación a la duración de los períodos de pre-oviposición y oviposición, los resultados obtenidos en el presente estudio son similares a los reportados para T. mexicanus criado sobre B. gasipaes, en donde se registraron 2,0 y 12,1 días, respectivamente (Stein y Daólio 2012). Sin embargo, la longevidad fue ligeramente superior (16,7 días). Contrariamente, Silva et al. (2009) registraron valores inferiores en la oviposición (6,21 días) y longevidad (8,83 días) de T. urticae criado sobre Gerbera jamesonii. Crooker (1985) estableció que la duración del período de preoviposición en tetraníquidos varía de 1 a 2 días, seguido de un período de oviposición, cuya duración depende de la especie de ácaro y de las condiciones ambientales, pero en promedio puede alcanzar entre 10 y 15 días.
El número total de huevos depositados por hembras de T. gloveri alcanzó sus máximos valores entre los 4 y 5 días, registrándose un promedio de 4,5 huevos hembra-1 día-1 (Figura 1). A partir de los 5 días se observó disminución de la tasa de oviposición hasta los días 11 y 12 de un promedio de 1,5 huevos hembra-1 día-1. Durante el día 13 se observó un ligero incremento en la oviposición, pero luego decreció hasta hacerse nula (Figura 1).
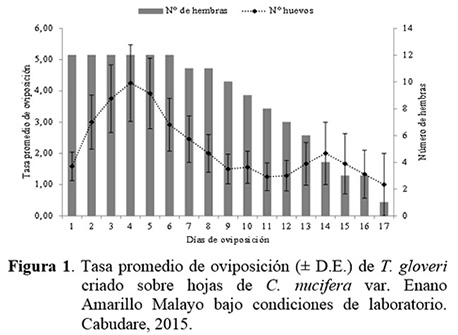
La fecundidad promedio de T. gloveri fue de 29,8 huevos hembra-1, con una tasa de oviposición de 2,9 huevos hembra-1 día-1 (Figura 1). Herbert (1981) reportó promedios de fecundidad para T. urticae de 37 huevos hembra-1, con valores máximos al tercer día de oviposición a 18ºC. Por otra parte, Giraud (1984) reportó promedios de fecundidad de 42,4 huevos por hembra, con una tasa de oviposición de 2,9 huevos hembra-1 día-1, con un valor máximo de 5,5 huevos hembra-1 día-1 alcanzado el cuarto día de oviposición. Las diferencias obtenidas en los valores de fecundidad de T. urticae podrían deberse a los cambios en el metabolismo de la planta hospedante, lo cual resulta en diferencias tanto en la tasa neta de reproducción (De Ponti 1977) como en los valores de rm, que dependen de la especie de la planta hospedante, la superficie disponible para cada individuo, la temperatura y la humedad (Gutierrez y Helle 1985).
Parámetros de la tabla de vida de T. gloveri
La tasa de supervivencia de T. gloveri fue máxima durante los primeros 5 días después de la emergencia de la hembra adulta, y luego comenzó a declinar hasta hacerse nula al día 12. La producción de progenie hembra hembra-1 día-1 (mx) fue relativamente constante durante casi todo el ensayo, excepto a los 7 días cuando se obtuvo el único ejemplar macho (Tabla 3, Figura 2). La tasa intrínseca de crecimiento (rm) del ácaro fue de 0,100 individuos hembra-1 día-1. La población se multiplicó (R0) 6,73 veces en un tiempo generacional (T) de 19,0 días. La tasa finita de crecimiento natural (λ) fue de 1,105 veces hembra día-1. Los resultados obtenidos para T. gloveri son superiores a los obtenidos para T. mexicanus criados sobre otras especies de palmera (B. gasipaes) en donde la tasa intrínseca de crecimiento (rm) del ácaro fue de 0,06 individuos hembra-1 día-1, la población se multiplicó (R0) 3,70 veces en un tiempo generacional (T) de 21,6 días y la tasa finita de crecimiento natural (λ) fue de 1,06 veces hembra día-1 (Stein y Daólio 2012). Así mismo, los valores de rm y λ obtenidos para T. gloveri son similares a los obtenidos para T. urticae en gerbera 0,12 para (rm) y 1,1 (λ) (Silva et al., 2009), pero inferiores a los obtenidos para Tetranychus ludeni (0,253 y 1,287) (Morros y Aponte 1994) y Tetranychus desertorum (0,188 y 1,207) (Tello et al. 2013) sobre hojas de caraota (Phaseolus vulgaris L.) y Tetranychus cinnabarinus (0,220 y 1,250) (Hazan et al. 1973). De acuerdo con Silva et al. (2009), los parámetros de la tabla de vida pudieron haber sido influenciados por la planta hospedera, lo cual sugiere la importancia del estudio de estos parámetros en especies de plantas que sirven como hospederas de T. gloveri. Adicionalmente, los valores de rm de los tetraníquidos puede ser mayor cuando el tiempo generacional es breve y la fertilidad es alta, puede también ser influenciada por superficie disponible para cada individuo y los valores de temperatura y humedad (Rivero y Vásquez 2009).
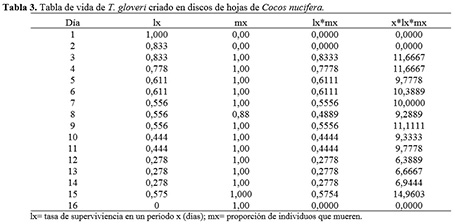
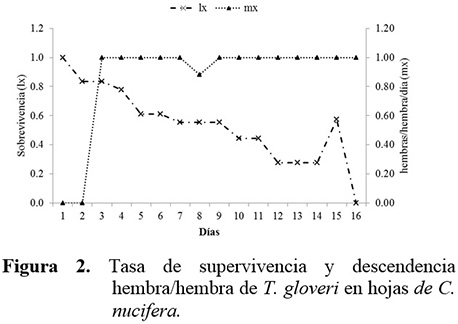
Los resultados de los parámetros de incremento poblacional obtenidos en el presente estudio indican que T. gloveri no expresó un alto potencial biótico al ser criado sobre hojas de C. nucifera, por lo que el daño económico causado sobre coco podría no ser importante bajo condiciones similares a las evaluadas en el presente estudio.
CONCLUSIONES
Los parámetros biológicos de T. gloveri, relacionados con el tiempo promedio de desarrollo (huevo-adulto), longevidad y fecundidad están dentro de los parámetros establecidos para otras especies relacionadas de Tetranychidae. Sin embargo, los valores de la tabla de vida permiten inferir que T. gloveri no expresa un alto potencial biótico sobre hojas de C. nucifera.
REFERENCIAS BIBLIOGRAFICAS
Adango, E; Onzo, A; Hanna, R; Atachi, P; James, B. 2006. Comparative demography of the spider mite, Tetranychus ludeni, on two host plants in West Africa. Journal of Insect Science 6:1-9. [ Links ]
Avery, DJ; Briggs, JB. 1968. The aetiology and development of damage in young fruit trees infested with fruit tree red spider mite, Panonychus ulmi (Koch). Annals of Appliet Biology 61(2):277-288. [ Links ]
Birch, L. 1948. The intrinsic rate of natural increase of an insect population. Journal of Animal Ecology 17(1):15-26. [ Links ]
Bolland, HR, Gutierrez J, Fletchmann, CHW. 1998. World catalogue of the spider mite Family (Acari: Tetranychidae).Linden, NL, Koninklijke Brill. 392 p. [ Links ]
Crooker, A. 1985. Embryonic and juvenile development. In Helle, W; Sabelis, MW. eds. Spider mites: their biology, natural enemies and control. Amsterdam, DE, Elservier. v. 1A, p. 149-163 [ Links ]
De Angelis, J; Berry, RE; Krantz, GW. 1983. Photosynthesis, leaf conductance, and leaf chlorophyll content in spider mite (Acari: Tetranychidae) injured peppermint leaves. Environmental Entomology 12(2):345-348. [ Links ]
De Ponti, OMB. 1977. Resistance in Cucumis sativus L. to Tetranychus urticae Koch: designing a reliable laboratory test for resistance based on aspects of the host-parasite relationship. Euphytica 26(3):641-654. [ Links ]
Fazlul-Hoque, M; Islam, W; Khalequzzaman, M. 2008. Life tables of two-spotted spider mites Tetranychus urticae (Acari: Tetranychidae) and its predator Phytoseiulus persimilis Athias-Henriot (Acari: Phytoseiidae). Journal Bioscience 16:1-10. [ Links ]
Flechtmann, CHW; Kreiter, S; Etienne, J; Moraes de, GJ. 1999. Plant mites (Acari) of the French Antilles. 1. Tetranychoidea (Prostigmata). Acarologia 40:137-144. [ Links ]
Gallardo, A; Vásquez, C; Morales, J; Gallardo, J. 2005. Biología y enemigos naturales de Tetranychus urticae en pimentón. Manejo Integrado de Plagas y Agroecología 74:34-40. [ Links ]
Gerson, U.; Weintraub, PG. 2012. Mites (Acari) as a factor in greenhouse management. Annual Review of Entomology 57:229-247. [ Links ]
Giraud, A. 1984. Biología, niveles poblacionales y control químico de Tetranychus cinnabarinus (Boisduval) y Tetranychus urticae Koch (Acari: Tetranychidae) en fresa. Tesis Mag. Maracay, VE, UCV. 127 p. [ Links ]
Gutierrez, J. 1985. Systematics. In Helle, W; Sabelis, M. eds. Spider Mites: their biology, natural enemies and control. Amsterdam, ND, Elsevier Science p. 75-90. [ Links ]
Gutierrez, J; Helle, W. 1985. Evolutionary Changes in the Tetranychidae. In Helle, W; Sabelis, M. eds. Spider Mites: their biology, natural enemies and control. Amsterdam, ND, Elsevier Science. p. 91-106. [ Links ]
Hazan, A; Gerson, U; Tahori, US. 1973. Life history and life tables of the carmine spider mite. Acarology 15(3):414-440. [ Links ]
Helle, W; Overmeer, W. 1985. Rearing Techniques. In Helle, W; Sabelis, M. eds. Spider Mites: their biology, natural enemies and control. Amsterdam, ND, Elsevier Science. p. 331-335. [ Links ]
Herbert, H. 1981. Biology, life tables and innate capacity for increase of the twospotted spider mite, Tetranychus urticae (Acarina: Tetranychidae). The Canadian Entomologist 113(5):371-378. [ Links ]
Migeon, A; Dorkeld, F. 2015. Spider mites web: a comprehensive database for the Tetranychidae (en línea). s.l. Consultado 2 jun. 2015. Disponible en http://www.montpellier.inra.fr/CBGP/spmweb. [ Links ]
Monjarás-Barrera JI; Lara-Villalón M; Juárez-Aragón MC; Torres-Castillo JA. 2015. New report of Tetranychus merganser Boudreaux and Oligonychus punicae Hirst on Moringa oleifera Lam. Southwestern Entomolgist 40(4):847-849. [ Links ]
Morros, ME; Aponte, O. 1994. Biología y tabla de vida de Tetranychus ludeni Zacher en caraota Phaseolus vulgaris L. Agronomía Tropical 44(4):667-677. [ Links ]
Ochoa, R; Aguilar, H; Vargas, C. 1994. Phytophagous mites of Central America: an illustrated guide. Turrialba, CR, CATIE. 234 p. [ Links ]
Rivero, E; Vásquez, C. 2009. Biologia e tabela de vida de Tetranychus desertorum (Acari: Tetranychidae) sobre folhas de feijão (Phaseolus vulgaris). Zoologia 26 (1):38-42. [ Links ]
Sances, FV; Wyman, JA; Ting, FP. 1979a. Physiological responses to spider mite infestations on strawberries. Environmental Entomology 8:711-714. [ Links ]
Sances, FV; Wyman, JA; Ting, FP. 1979b. Morphological responses of strawberry leaves to infestations of twospotted spider mite. Environmental Entomology 72:710-713. [ Links ]
Silva, EA; Reis, PR; Carvalho, TMB; Altoé, BF. 2009. Tetranychus urticae (Acari: Tetranychidae) on Gerbera jamesonii Bolus and Hook (Asteraceae). Brazilian Journal Biology 69(4):1121-1125. [ Links ]
Stein, CP; Daólio, N. 2012. Biologia de Tetranychus mexicanus (McGregor) (Acari: Tetranychidae) em folhas de pupunha (Bactris gasipaes Kunth). Bioikos 26(1):23-28. [ Links ]
Tanigoshi, LK; Davis, RW. 1978. An ultrastructural study of Tetranychus mcdanieli feeding injury to the leaves of Red Delicious apple (Acari: Tetranychidae). International Journal of Acarology 4:47-56. [ Links ]
Tello, V; Castillo, P; Briceño, R; Sánchez, M. 2013. Parámetros biológicos de Tetranychus desertorum (Acari: Tetranychidae) sobre hojas de poroto. Idesia 31(4):27-33. [ Links ]
Tomczyk, A; Kropczynska, D. 1985. Effects of the plants. In Helle, W; Sabelis, MW. eds. Spider mites: their biology, natural enemies and control. Amsterdam, ND, Elsevier Science. v. 1A, p. 317-329 [ Links ]
Vásquez, C; Colmenárez, M; Valera, N; Díaz, L. 2007. Antibiosis of kidney beans cultivars to the carmine spider mite, Tetranychus cinnabarinus (Boisduval) (Acari: Tetranychidae). Integrated Control of Plant-feeding Mites. IOBC/wprs Bulletin 30(5):133-138. [ Links ]
Vrie van de, M; McMurtry, JA; Huffaker, CB. 1972. Ecology of tetranychid mites and their enemies - a review - III: biology, ecology, and pest status, and host-plant relations of tetranychids. Hilgardia 41(13):343-432. [ Links ]
Wrensch, DL. 1985. Reproductive parameters. In Helle, W; Sabelis, M. eds. Spider mites: their biology, natural enemies and control. Amsterdam, ND, Elsevier Science. p. 165-168. [ Links ]