









 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
Permalink
INTRODUCCIÓN
La malaria es una enfermedad transmitida a los humanos a través de una picadura de un mosquito hembra del género Anopheles infectado con parásitos del género Plasmodium Se estima que, en 2010, hubo 216 millones de casos de malaria en el mundo y que esta enfermedad causó más de 150,000 muertes1.
Desde 2003, el control de la malaria se ha considerado exitoso, con solo 91 casos en 2009 y 23 casos entre enero y julio de 2010 (16 de ellos fueron casos autóctonos). Entre los casos, 94% eran del departamento de Alto Paraná y 6% eran del departamento de Caaguazú2.
Actualmente, el Servicio Nacional para la eliminación de la enfermedad (SENEPA) considera los departamentos de Alto Paraná y Caaguazú, que fueron regiones históricamente endémicas, como regiones con bajo riesgo de transmisión (2).
La resistencia a insecticidas es uno de los ejemplos más interesantes de la rapidez con la que se dan los cambios a nivel evolutivo. El DDT y los piretroides son neurotoxinas que actúan sobre los canales de sodio dependientes de potencial modificando la cinética de su abertura, lo que resulta en una abertura prolongada de cada canal lo que conlleva a la parálisis y muerte del insecto. Uno de los mecanismos de resistencia a piretroides en insectos se conoce como resistencia knockdown (KDR en inglés) causado por la disminución de la sensibilidad en ese canal3.
Los piretroides son los insecticidas más comúnmente usados para fumigación residual de interiores y el único en su clase, recomendado para la impregnación de mosquiteros debido a su relativa baja toxicidad para los mamíferos, pero con capacidad de producir efectos knockdown en insectos4.
Se ha investigado sobre la variabilidad del ADN en el gen del canal de sodio dependiente del potencial, comúnmente referido como gen VGSC (siglas en ingles por voltaje gatedsodium cannel), que codifica los segmentos 5 y 6 del dominio II transmembranal de la proteína, de manera a estimar la evolución de la resistencia a piretroides en poblaciones de manzanas a nivel mundial. La variabilidad del ADN en 38 secuencias, revelaron la existencia de una mutación KDR única (L1014F), involucrada en la resistencia a piretroides y se encuentra en esta región mencionada, la cual es el resultado de varias sustituciones convergentes5.
El fenotipo de Resistencia en el gen VGSC es comúnmente referido por una mutación particular (L1014F/S/H) en el segmento IIS66,7. Otras mutaciones en diferentes regiones del gen también confieren resistencia del tipo knockdown en algunos insectos6,8, pero entre los anophelinos este es el único locus donde las mutaciones han sido reportadas hasta ahora y que confieren resistencia9-15.
La presencia de la mutación,así como también sus efectos, han sido plenamente estudiadas en mosquitos anopheles. Hasta ahora, las mutaciones KDR han sido ya detectadas en al menos 13 especies (Anopheles gambiae, Anopheles arabiensis, Anopheles sinensis, Anopheles stephensi, Anopheles subpictus, Anopheles sacharovi, Anopheles culicifacies, Anopheles sundaicus, Anopheles aconitus, Anopheles vagus, Anopheles paraliae, Anopheles peditaeniatus and Anopheles albimanus) de poblaciones africanas, asiáticas y recientemente del continente americano. Se han descrito siete variantes de la mutación (L1014F, L1014S, L1014C, L1014W, N1013S, N1575Y y V1010L) pero con una alta prevalencia de la mutación L1014F16.
El objetivo del presente trabajo consistió en detectar y caracterizar mutaciones KDR por técnicas moleculares en vectores anophelinos capturados en localidades históricamente endémicas bajo vigilancia epidemiológica.
MATERIALES Y MÉTODOS
Se realizó un estudio observacional descriptivo de corte transversal empleando muestras de mosquitos Anoheles de las especies A. strodei, A. albitarsis, A. fluminensis, A. evansae, A. nuneztovari, A. nyssorhynchela lutzi y A. oswaldoi capturados en criaderos naturales, en los departamentos de Caaguazú y Alto Paraná en Paraguay, durante los meses de agosto, setiembre y noviembre de 2016 y diciembre de 2017. Estos meses se caracterizan por temperaturas cálidas y corresponden a la estación primaveral del país.
Los estudios fueron realizados en pooles ya que estas muestras se utilizaron previamente para otro proyecto y ya quedó el material genético guardado a -20 grados. Muestras disponibles: A. strodei (17 pooles), A. albitarsis (4), A. fluminensis(1), A. evansae(1), A. nuneztovari(1), A. nyssorhynchela lutzi(1) y A. oswaldoi(1). Número máximo de mosquito por pool fue de 10 y mínimo de 3.
Los pooles de mosquitos fueron sometidos a extracción de ADN empleando el kit Nucleo Spin® DNA Insect de Macherey-Nagel GmbH& Co siguiendo el protocolo del mismo previo almacenamiento.
Para la identificación de la mutación KDR se empleó la técnica PCR anidada, siguiendo el protocolo propuesto por Lol J Cet al 2013. Empleando los cebadores AAKDRR (5′-GCAANGCTAAGAANAGRTTNAG-′3); AAKDRF (5′-AGATGGAAYTTYACNGAYTTC-′3) y AAKDRF2 (5′-CATTCATTTATGATTGTGTTTCGTG-′3)(17).La amplificación del segmento 6 del dominio II del gen VGSC empleando cebadores reportados se llevó a cabo en una mezcla de reacción de 25 µL que contenía 17.55µL de agua. 2.5 µL de Buffer (10x), 2 µL MgCl2 (25x), 0.5 µL de dN TPs, 0.1 µL de cada cebador (AAKDRR y AAKDRF), 0.25 µL de Polimerasa Hot Start Taq (Thermo scientific) y 2 µL de ADN. Las condiciones de PCR consistieron en una desnaturalización inicial a 95º C por 3 min., seguido de 35 ciclos de 95º C por 45 sec., 40.5º C por 45 sec. y 72º C por 1 min. con una elongación final a 72º C por 5 min en un termociclador Simpli AmpTMThermo Fisher Scientific. Se realizó una segunda PCR empleando los cebadores AAKDRR y AAKDRF2 utilizando las mismas especificaciones de reacción que en la primera PCR y como ADN molde el producto de la primera PCR. Las condiciones de la segunda PCR consistieron en una desnaturalización inicial a 95º C por 3 min., seguido de 40 ciclos de 95º C por 45 sec., 51.5º C por 45 sec. y 72º C por 1 min. con una elongación final a 72º C por 5 min en un termociclador Simpli AmpTMThermo Fisher Scientific.
La muestra fue secuenciada empleando el servicio de secuenciación de Macrogen® y la secuencia fue analizada empleando el software en línea Clustal Omega®
RESULTADOS
Amplificación de KDR
Todos los pooles de cada especie fueron analizados a excepción de los pooles de A. strodei que se analizaron 9 de 17.
Los cebadores empleados fueron específicos para un alelo de kdr en el segmento 6 del dominio II del gen VGSC en mosquitos de la especie Anopheles strodei, no así para las demás especies de Anopheles empleadas para el ensayo en la cual no hubo amplificación.
Secuenciación
Los resultados de la secuenciación de una muestra de pool de la especie An. Strodei confirmó la amplificación del segmento del gen VGSC, con la presencia del tipo susceptible (wild type) TTA (L1014) (Figura 1).

Figura 1: Alineación del segmento del gen VGSC de An. strodeide Paraguay con An. albimanus de América latina (Colombia susceptible TTG L1014, Costa Rica resistente TTC 1014F y Mexico resistente TGT 1014C). El sitio polimórfico (posición aminoacídica 1014) está encerrado por un cuadro rojo.
Los picos en el electroferograma son únicos para el sitio de interés analizado, lo que demuestra la uniformidad del polimorfismo observado en cada mosquito del pool (Figura 2).
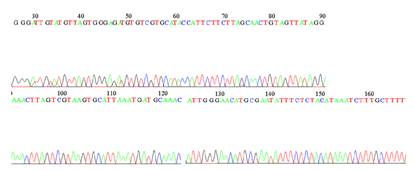
DISCUSIÓN
La caracterización de las mutaciones KDR en especies anophelinas de Latinoamérica todavía es bastante difícil debido a que no existen secuencias del gen VGSC completas o solo existen secuencias parciales disponibles en línea. La mayoría de las secuencias disponibles son de mosquitos de interés vectorial para la transmisión de la Malaria en África.
Los primers reportados para A. albimanus de Guatemala han demostrado ser específicos para amplificar ADN de A. strodei de Paraguay17. Si bien con esto pudimos obtener el producto de PCR del gen VGSC como para secuenciar y observar la versión susceptible del polimorfismo L1014, aun debemos colectar más mosquitos de la misma especie y realizar el monitoreo para conocer la frecuencia de es este polimorfismo. Es importante tener en cuenta que el Anopheles darlingi realmente es el que está considerado como el vector primario y el Anopheles albitarsis como vector secundario en nuestro país. En este contexto, las muestras empleadas en este estudio provienen de un estudio anterior donde la colecta se realizó en regiones históricamente endémicas de Paraguay (Caaguazú y Alto Paraná) y donde se colecto gran cantidad de la especie A. strodei en comparación con las otras especies anophelinas estudiadas, A. albitarsis, A. fluminensis, A. evansae, A nuneztovari, A.nyssorhynchela lutzi y A.oswaldoilo que podría ser de interés para la vigilancia entomológica.
Muy pocas especies de anopheles de Latinoamérica han sido caracterizadas en su polimorfismo L1014 para el gen VGSC, por tanto, existe una carencia de secuencias disponibles o métodos moleculares que permitan estudiar a los mosquitos Anopheles locales, constituye por tanto, un desafío establecer la técnica de PCR con los cebadores apropiados.
La secuenciación de la especie Anopheles strodei mostró que el pool analizado contenía el alelo kdr susceptible (wild type) TTA (L1014), previamente reportado en An. sacharovi, An. sinensis, An. albimanus y otras especies de Anopheles16,17,18,19,20. En otras poblaciones de mosquitos en Latinoamérica se ha detectado una mutación homocigota no sinónima, TGT (cisteína, L1014C), en muestras de campo de México. Esta mutación está asociada con la resistencia a permetrina, deltametrina y beta-cipermetrina en Ansinensis16,21,22. También se ha reportado en una muestra de Costa Rica contenía un polimorfismo TTC homocigota (fenilalanina, 1014L), esta mutación se asocia a la resistencia a permetrina y DDT en An. gambiae, resistencia deltametrina en Ansinensisy resistencia a DDT, permetrina, alfa-cipermetrina, lambda-cihalotrina y etofenprox en Anpeditaeniatus9,19,22.
Si bien analizamos 9 de 17 pooles de A. strodei porque consideramos suficiente para probar que los cebadores eran específicos, desde el punto de vista entomológico consideramos importante mencionar la existencia de los 17 pooles de dicha especie porque corresponden a un estudio previo, donde se puede observar que de la colecta realizada en departamentos históricamente endémicos, la especie mayormente colectada es el A. strodei, y podría constituirse en un potencial vector secundario de interés.
Hasta donde sabemos, ésta es la primera caracterización del gen VGSC en mosquitos Anopheles del Paraguay y para la especie A. strodei. Disponer de técnicas moleculares que permitan amplificar este gen, es importante para el monitoreo de sitios polimórficos que están asociados a conferir resistencia a insecticidas y sería un apoyo importante en la vigilancia para mantener al país libre de Malaria.