











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink
INTRODUCCIÓN
La ganadería extensiva está asociada a la deforestación por el cambio de uso del suelo de bosques a pastos, pero también es un motor directo de degradación de los bosques al aumentar la matriz de pastos, generando un borde más abrupto entre los relictos boscosos y los pastizales adyacentes quedando así los bosques expuestos al pastoreo constante (Meza Elizalde y Armenteras Pascual, 2018).
En el caso de los bosques Amazónicos, la colonización agropecuaria y la praderización de áreas forestales, es sin duda, el tipo de intervención antrópica que mayor impacto genera (Murcia et al., 2014), y es impulsada por patrones indirectos como fuerzas de mercado, pobreza, poca vocación y desconocimiento de tenencia de las tierras (Kaimowitz y Angelsen, 2001).
En Colombia, la ganadería es la actividad más representativa del sector agropecuario, predominando en el territorio nacional en 27 de los 32 departamentos, y siendo nueve veces mayor el área dedicada a esta actividad sobre el área agrícola (Vergara, 2010). De acuerdo a la encuesta nacional agropecuaria (Departamento Administrativo Nacional de Estadística [DANE], 2017), el 78,2% de la superficie de uso del suelo se dedica a la actividad pecuaria, 7,3% en sistemas agrícolas y 2,5% en otros usos, mientras que tan solo un 11,9% se mantiene en bosques. Así mismo, el 70% de la ganadería bovina del país se encuentra bajo sistemas de producción extensivos con una capacidad de carga promedio de 0,9 animales por hectárea (Mahecha et al., 2002).
La deforestación a causa de la conversión de bosques a áreas agropecuarias es mayor en la región Amazonia con el 46,9% del total nacional y se concentra en los departamentos de Guaviare, Caquetá y Putumayo (Instituto de Hidrología, Meteorología y Estudios Ambientales [IDEAM], 2019). Siendo el departamento del Guaviare uno de los focos de deforestación con cerca del 23% de su área intervenida (Niño, 2010), con una cobertura de 1.548.75 km2 de bosques fragmentados, 4.574.2 km2 en pastizales, y con una tasa media de praderización anual de 298,8 km2/año (Murcia et al., 2014). En el Guaviare, el municipio de El Retorno es considerado el epicentro ganadero del departamento y el polo de desarrollo para esta actividad. Constituyéndose en uno de los municipios que presenta mayor tasa de deforestación anual (6492,65 Ha/año) para el establecimiento de pastos (Murcia et al., 2014).
La ganadería en este municipio se ha establecido bajo un sistema extensivo principalmente para la producción de carne (Ramírez, 2009), teniendo una participación del 1% en la producción nacional con aproximadamente 264.300 cabezas de ganado, pertenecientes en su gran mayoría a pequeños y medianos productores (Bermúdez, 2016). La ganadería en el municipio tiene un crecimiento en el mercado fluctuante, ya que en determinadas épocas del año se presenta sobre-oferta de algunos productos con la consecuente pérdida del producto, trayendo consigo deterioro de los ingresos y en consecuencia, de la calidad de vida de los productores (Ecoservig, 2009).
La actividad ganadera que se desarrolla es de carácter estabulado, extensivo y está asociada al establecimiento de gramíneas, la tala y quemas incontroladas (Mahecha et al., 2002). Este modelo productivo extensivo ha generado múltiples conflictos socio-ecológicos en el territorio (Arias-Patiño, Hurtado-Delgado, Gualaco-Morales y Jiménez-Ávila, 2015), ya que no solo genera la perdida de área y fragmentación de los bosques (Foley et al., 2005), sino que también los expone a degradación por otros impactos directos propios de la actividad ganadera. Afectando, por ejemplo, el recurso hídrico, dado que es común el uso de fuentes hídricas naturales al interior de los bosques como bebederos para el ganado (Murcia et al., 2010). El manejo del ganado dentro de las áreas forestales constituye uno de los principales determinantes de su diversidad florística (Milchunas et al., 1988). Esta actividad, no solo afecta el número de especies presentes sino también la abundancia y composición específica de las comunidades (Pykala, 2005; Stockton et al., 2005; Altesor, Piñeiro, Lezama, Jackson y Sarasol, 2006), favoreciendo el crecimiento de ciertos tipos funcionales de plantas por sobre otros (Lavorel, Mcintyre y Grigulis, 1999; Díaz, Lavorel, Mcintyre, Falczuk y Casanoves, 2007) y modificando el balance entre la colonización y extinción localizada de especies (Olff y Ritchie, 1998).
El pastoreo en bosques también hace posible el establecimiento de gramíneas, afectando de forma negativa la viabilidad de poblaciones forestales a futuro (Tomimatsu y Ohara, 2003). Aunque, claro está, influyen otros factores como la disponibilidad y diversidad de propágulos en la regeneración natural, junto a la disponibilidad de sitios seguros para el establecimiento de plántulas (Loydi y Distel, 2010). Dado que estos cambios se pueden registrar en la estructura horizontal y vertical de la vegetación y son derivados de la distancia que recorren los bovinos para buscar agua dentro del bosque (Morici et al., 2003), el objetivo del presente estudio es comparar y analizar las variaciones de los atributos estructurales, florísticos y de diversidad de la regeneración natural, comparando áreas boscosas con presión ganadera y áreas que están adyacentes a cobertura de pastos sin uso y a cultivos de caucho.
MATERIALES Y MÉTODOS
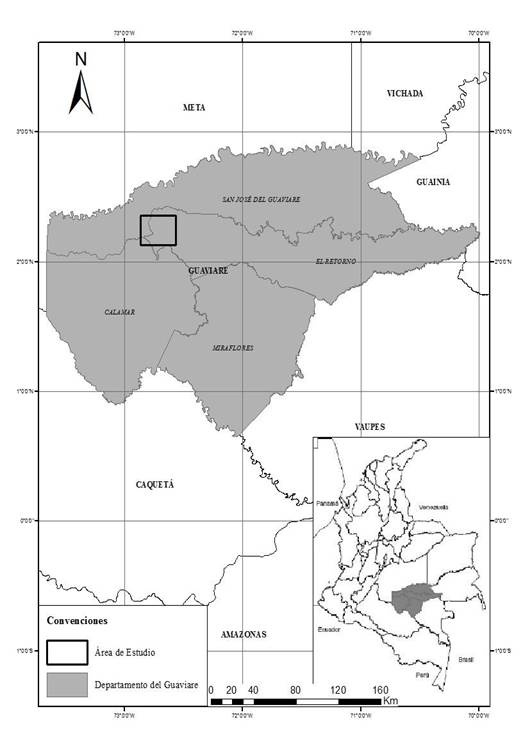
Área de Estudio: El estudio fue realizado en el norte de la Amazonia Colombiana, específicamente en el departamento del Guaviare en el municipio El Retorno. Este municipio, se encuentra ubicado en la cuenca alta del Río Inírida, a una altitud de 420 m.s.n.m., temperatura promedio de 25 °C, precipitación anual entre 2000 y 4000 mm, humedad relativa del 85 % (Instituto Geográfico Agustín Codazzi [IGAC], 2008), correspondiente a la zona de vida de bosque húmedo tropical (bh-T). Este departamento tiene una vocación ganadera representada en un 7,3% lo cual ha generado una transformación de coberturas naturales como los bosques en pastizales homogéneos con menos de 30 especies de plantas nativas (Bautista, 2020; Etter, 1990) (Figura 1).
Métodos:
Diseño de muestreo: Se seleccionaron relictos boscosos que no han presentado extracción de productos forestales en los últimos 30 años, los cuales limitan con pastos y plantaciones de caucho, estas zonas fueron denominadas “ambientes contrastantes”.
El primer ambiente contrastante (AC1) corresponde al relicto de bosque adyacente a áreas praderizadas con la especie Olyra latifolia pero que no tiene uso ganadero. El segundo ambiente contrastante (AC2) es un bosque contiguo a áreas dedicadas a la producción ganadera extensiva doble propósito de bovinos por pequeños productores, los cuales permiten la libre circulación de los animales al interior del bosque para su alimentación e hidratación. Estos dos ambientes contrastantes sufren de quemas inducidas por finqueros, que tienen como fin hacer limpias para expandir su área productiva, lo que trae como consecuencia la expansión de la frontera agropecuaria en la zona. Por último, se evalúo como tercer ambiente contrastante (AC3), un área de bosque adyacente a áreas agrícolas de plantaciones permanentes de caucho con edades variables (3 a 18 años en producción), perteneciente a productores asociados a la asociación de caucheros -ASOPROCAUCHO-.
En cada ambiente contrastante, se establecieron tres (3) transectos desde el borde de la cobertura adyacente hacia el interior del bosque, de un ancho fijo de 4 metros y una longitud de 100 m para la medición de todos los latizales (DAP <10 cm; Altura total >1,5 m). A lo largo de cada transecto se establecieron parcelas cuadradas de 4 m2, separadas entre sí 10 m, en las cuales se midieron brinzales (DAP < 10 cm; HT > 0,3 m), y estas parcelas a su vez se subdividieron en parcelas de 1 m2 para hacer el conteo de renuevos (DAP < 10 cm; HT < 0,3 m).
El listado de composición florística del área de estudio estuvo conformado por todas las especies vegetales de hábito arbóreo, arbustivo, herbáceo, lianas y palmas. Por el contrario, en el análisis estructural solo se tuvieron en cuenta las especies de hábito arbóreo. El material botánico que se colecto en todas las parcelas fue procesado en el Herbario Forestal Gilberto Emilio Mahecha Vega de la Universidad Distrital Francisco José de Caldas (UDBC). Para la determinación taxonómica del material vegetal se recurrió a claves taxonómicas y comparación directa con los ejemplares del herbario.
Análisis de datos:
Diversidad y Riqueza de la regeneración natural
Para efectos del presente estudio se consideró como diversidad a la riqueza de especies o la distribución proporcional de su abundancia (Whittaker, 1973) y riqueza como el número de taxa que constituyen un área particular (Rangel y Velásquez, 1997).
Se determinó la composición y estructura de la regeneración natural en cada uno de los ambientes contrastantes, para lo cual se calculó el Índice de Regeneración Natural (RN) para cada especie forestal a partir de la media aritmética de los valores de Abundancia, Frecuencia y Clase de Tamaño.
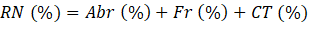
Dónde:
Abr (%): Abundancia relativa de las especies en la regeneración natural (latizales, brinzales y renuevos)
Fr (%): Frecuencia relativa de las especies en la regeneración natural (latizales, brinzales y renuevos)
CT (%): Clase de tamaño
Siendo la clase de tamaño, una medida útil basada en la altura y diámetro de los individuos que se emplea para conocer la estratificación de la comunidad (Rangel y Garzón, 1994).
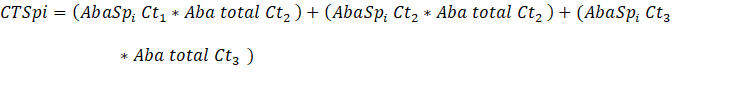
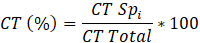
Dónde:
Aba: Abundancia absoluta (Número de individuos por clase de tamaño)
Ct1: Categoría de tamaño latizal
Ct2: Categoría de tamaño brinzal
Ct3: Categoría de tamaño renuevo
Para el análisis florístico, se tuvieron en cuenta las especies con hábitos herbáceos, arbustivos, arbóreos, lianas y palmas. Este trabajo toma los números efectivos de la diversidad verdadera de Jost como medidas de diversidad, incluyendo datos de abundancia relativa de diferentes especies vegetales en tres ambientes contrastantes. La estimación de la riqueza para todas las especies encontradas en los tres ambientes se realizó a partir del cálculo de tres órdenes de riqueza (q0, q1, q2), representados por el número de especies por tratamiento (q0), el inverso de Shannon (q1), el inverso de Simpson (q2), con intervalos de confianza al 95%, procesados en el paquete estadístico iNEXT (Hsieh et al., 2014) en el programa estadístico versión 3.6.3 R.
Para la evaluación de la eficiencia de muestreo se construyeron curvas de rarefacción usando la abundancia de todos los individuos muestreados por cada ambiente contrastante AC1-AC2-AC3; para calcular la rarefacción se usó el paquete iNEXT (Hsieh et al., 2014).
Por último, se empleó el análisis de correspondencia múltiple para las variables cualitativas de ambiente contrastante, hábito, gremio, especies con mayor porcentaje de regeneración por ambiente contrastante. Esto determinó el porcentaje de varianza y posible ordenación de los resultados en el presente estudio. Para el análisis de estos valores se usó la librería FactoClass.
Como una aproximación al estado sucesional de las áreas evaluadas, empleamos los gremios ecológicos. Estos gremios son un indicador de la dinámica del bosque asociada a los relevos florísticos en el tiempo en relación a la forma cómo las plantas usan los recursos, (Whitmore, 1990). Los gremios ecológicos se pueden clasificar según los requerimientos de luz en (Finegan, 1996; Linares, 2000): 1) Heliófitas efímeras o pioneras, son aquellas especies que requieren plena insolación durante toda su vida, de crecimiento rápido y vida corta, e importantes en la colonización de sitios abiertos, además de mantener el banco de semillas del suelo. 2) Heliófitas durables o secundarias tardías, son especies intolerantes a la sombra, de crecimiento rápido y ciclo de vida más bien largo. Se apropian de sitios de claros después que las heliófitas efímeras desaparecen, estableciéndose por un período de tiempo mucho más largo. 3) Esciofitas parciales, que corresponden a las plantas que son tolerantes a la sombra en las etapas tempranas de su desarrollo, aunque requieren de iluminación para pasar de las etapas intermedias a la madurez, soportan la sombra, pero no exclusivamente y aumentan el crecimiento si se abre el dosel del bosque, permitiendo mayor ingreso de luz solar. Por último, 4) Esciófitas totales, que son especies que requieren solamente sombra, y en el tiempo no tienen la capacidad de aumentar significativamente su crecimiento con la apertura del dosel. (Finegan, 1996; Linares, 2000).
RESULTADOS Y DISCUSIÓN
Diversidad y Riqueza de la regeneración natural
En general se registraron 281 especies distribuidas en 63 familias botánicas (Anexos). Las curvas de rarefacción para los tres ambientes contrastantes presentan una asíntota horizontal para todos, determinando así que el muestreo fue adecuado teniendo en cuenta la rarefacción. El AC3 registró mayor diversidad y menor riqueza, siendo éste quien difiere significativamente en términos de riqueza. En el AC2 se observó menor diversidad y menor riqueza en comparación con los otros dos ambientes, mientras que en el AC1 hubo una mayor riqueza, pero menor diversidad.
La rarefacción estimada a partir del número de individuos muestreados, determinó que el muestreo fue el adecuado para la zona de estudio. Por otro lado, se observa que el número de especies difiere en los tres ambientes contrastantes, encontrando que, a partir de una muestra de 367 individuos, el número de especies para el AC1 es (139) especies, para AC2 es (106) especies y para el AC3 (170) especies. El AC1 y AC2 tienen similitud ya que ambas tienen menor diversidad en comparación a AC3 (Figura 2).
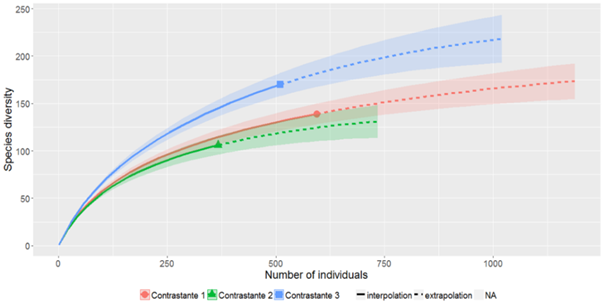
Figura 2. Curva de acumulación de especies (Riqueza) en la regeneración natural por rarefacción en los tres ambientes contrastantes.
En la (Figura 3) se presentan los resultados de diversidad verdadera de Jost para ordenes de (q=0, q=1 y q=2). La riqueza estimada a partir del número efectivo de especies presentó una completitud del 94,5% para los tres ambientes, bajo un intervalo de confianza estadístico del 95% para el muestreo.
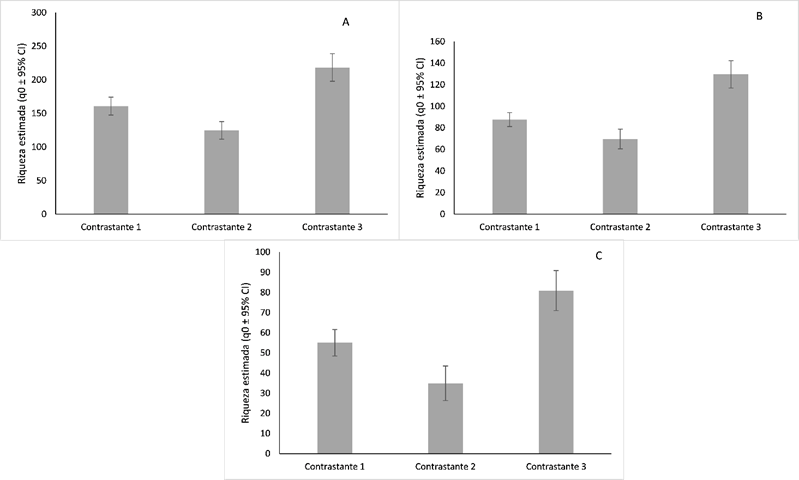
Figura 3. Índices de Diversidad Verdadera (riqueza estimada a partir del número efectivo de especies) para la regeneración natural en los 3 ambientes contrastantes.
Considerando la riqueza de especies observada y la riqueza esperada (q=0) se pudo calcular qué tan completo fue el inventario de los sitios para representar la máxima riqueza potencial de acuerdo con el muestreo, indicando que el AC3 (Bosque adyacente a caucho) es 26.1% inferior en riqueza de especies respecto al AC1 y 43,1% al AC2. El AC1 comprendió 160,8 especies efectivas, AC2: 124,7 especies efectivas y AC3: 218,3 número efectivo de especies. Por lo tanto, el AC3 es superior en diversidad respecto a los AC1 y AC2.
En el segundo orden de riqueza inverso de Shannon (q1) estimado a partir de la abundancia, se observó una repuesta similar. El AC2 (bosques adyacentes a pastos con ganado) fue significativamente inferior en abundancia a los ambientes de bosques lindantes a pastos sin uso (AC1: 20,4%) y bosques adyacentes a caucho (AC3: 46,2%). Esta respuesta puede ser atribuida a la mayor abundancia de especies como Guatteria cf. metensis, Petrea volubilis, Ocotea Sp. 4, Inga acreana, Sterculia apetala, Neea floribunda, Brosimum guianense, Sorocea steinbachii, Protium heptaphyllum, Inga brachyrhachis, Ing cf. heterophylla, Siparuna decipiens. Este orden de riqueza (q=1) reveló un número efectivo de especies de AC1:87,5, AC2:69,7 y AC3: 129,5. Para el último orden de riqueza (q2), presenta un mayor número de especies dominantes (altos valores de abundancia y comunes en los ambientes). El AC3 con numero efectivo de especies de (80,8), en comparación al AC1 (36,6%) y numero efectivo de especies de (55,05) y AC2 (56,7%) y (80,8%) número efectivo de especies, debido a la presencia de especies como Guatteria cf. metensis, Himatanthus articulatus, Inga acreana, Brosimum guianense y Sorocea steinbachii.
Estructura de la Regeneración Natural
En los bosques adyacentes a pastos sin uso (AC1), se registraron 140 especies forestales distribuidas en 44 familias botánicas, siendo las familias más representativas LEGUMINOSAE con 11 especies, MORACEAE con 10 especies y MELIACEAE con 9 especies. En las áreas de bosque con pastoreo (AC2), se registraron 108 especies pertenecientes a 42 familias, donde las familias con mayor cantidad de especies fueron LEGUMINOSAE con 12 especies, BIGNONIACEAE con 7 especies y en tercer lugar MELASTOMATACEAE y BURSERACEAE con 6 especies, respectivamente. Por último, en las áreas de bosque adyacentes a cultivos permanentes de caucho, se obtuvo un registro de 172 especies distribuidas en 47 familias, siendo la familia LEGUMINOSAE con 17 especies la más representativa, seguida de LAURACEAE y MORACEAE con 14 especies cada una. Las demás familias de las tres áreas muestreadas tienen una representación de 9 a 1 especies cada una.
Las especies que presentaron mayor valor de Índice de Regeneración Natural (RN %) debido a sus valores de abundancia, frecuencia y clase de tamaño, en AC1 fueron, Miconia truncata (4,2%), Protium sagotianum (3,4%) y Virola peruviana (3%); para AC2, la especie Triplaris americana (15,59%) obtuvo el mayor índice de regeneración, seguido de Otoba sp1 (4,3%) y Miconia elata (4%), y en AC3, las especies Guatteria cf. metensis (4,3%), Himatanthus articulatus (3,7%) e Inga acreana (3,3%) representaron el mayor índice de regeneración (Figura 4).
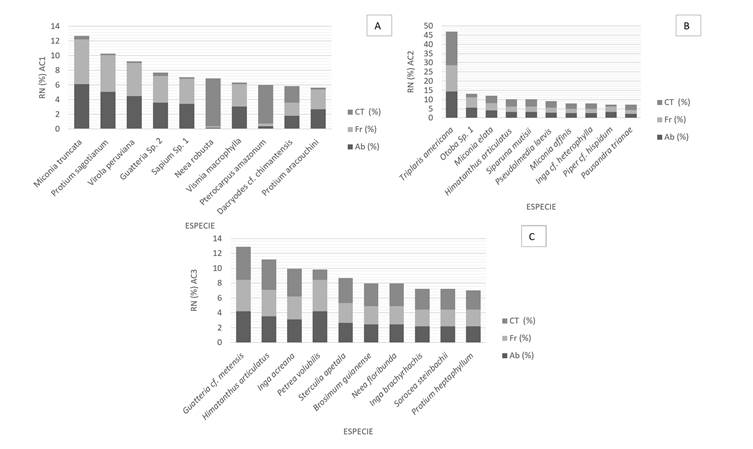
Figura 4. Índice de Regeneración natural para los tres ambientes contrastantes AC1 (A), AC2 (B) y AC3 (C).
Teniendo en cuento los resultados obtenemos que el bosque adyacente a áreas con cultivo de caucho AC3 cuenta con la mayor diferencia real de diversidad y en número efectivo de especies, en comparación con el bosque adyacente a pastos sin uso, y obtenemos la menor diversidad en las áreas de bosques con entrada de ganado.
En general, se comprueba que las especies heliófitas son las más abundantes en tres ambientes contrastantes, siendo afín a lo obtenido por Lescano et al., 2002 para fragmentos de áreas muy pequeñas (< 10 ha) de bosques. Así mismo, se confirma que el incremento de la regeneración natural es una respuesta a las intervenciones antrópicas (pastoreo) y se evidencia en la presencia de especies clasificadas como heliófitas durables (Graaf, Poels y Van Rompaey, 1999; Rose, 2000; Arets, Van der Hout y Zagt, 2003; Peña et al., 2008).
Se evidenció que en los bosques adyacentes a plantaciones de caucho (AC3), la regeneración natural tiene mejores atributos estructurales, florísticos y de diversidad, en comparación a los bosques en los cuales entra el ganado (AC2). Lo anterior permite inferir que hay una degradación de los bosques con pastoreo y adyacentes a cobertura de pastos, en los cuales hay una mayor abundancia y frecuencia de especies herbáceas que colonizan ambientes perturbados.
La degradación de los bosques se relaciona directamente con la carga animal, ya que la diversidad y configuración estructural de especies en nuestro estudio disminuyo, conforme a la cercanía con la cobertura de pastos. Lo anterior, está directamente relacionado con la carga animal en las áreas adyacente a los bosques y por tanto su transformación a pastos y configuración ecológica actual, ya que al año del desarrollo de la presente investigación se resalta el aumento de cabezas de ganado con 281.611 cabezas en 2016 a 405.188 en 2017 en el departamento (Bautista, 2020).
La diversidad en los ambientes contrastantes fueron diferentes entre sí. Se encontró que el área de menor diversidad fue el ambiente de bosques con pastoreo, tanto en número de especies (q0), abundancia (q1), y dominancia (q2), demostrando que características estructurales y de composición, se vea afectadas por la entrada del ganado en busca de alimento y agua.
En las áreas donde hay pastoreo se registran especies de habito herbáceo y lianas, como Pyrostegia dichotoma, Bignonia aequinoctialis, Solanum jamaicense y Bauhinia tarapotensis, las cuales aparecen en los sitios abiertos por el paso del ganado durante su traslado del borde al cuerpo de agua. Lo anterior es soportado por Oesterheld y Sala (1990), quienes exponen que la herbívora crea sitios favorables para la germinación e instalación de nuevos individuos y hábitos y el aumento de especies forrajeras en las áreas usadas como abrevadero dentro del bosque.
La menor diversidad observada en el ambiente de pastoreo también se podría atribuir al pisoteo continuo, lo cual dificulta el establecimiento y supervivencia de las plántulas (Brits, Van Rooyen y Van Rooyen, 2002). Estos comportamientos negativos han sido observados en otros estudios de Milchunas et al. (1988) y Estelrich, Chirino, Fernández y Morici (1997). La baja diversidad en bosques adyacentes a pastos, concuerda con lo encontrado por Meza Elizalde y Armenteras Pascual (2018) quienes indicaron que en los bosques que limitan con pastos hay mayores valores de dominancia y menores valores de equidad de acuerdo al valor obtenido en el Índice de Shannon.
El área circundante a bosques de caucho presentó la mayor diversidad entre las tres zonas, por ende, se asume que el caucho adyacente al bosque, proporciona condiciones que facilitan el reclutamiento de especies de estados seriales más avanzados en contraste con zonas que reciben una mayor exposición solar y al viento en el borde (áreas adyacentes a pastos). De acuerdo a Meza Elizalde y Armenteras Pascual (2018), las plantaciones de caucho tienen la capacidad de amortiguar el microclima del bosque adyacente, proporcionando condiciones más frías y húmedas con menor presión de vapor cerca de los bordes. Baker et al., (2014) y Wuyts et al. (2008), indican que coberturas de porte alto al lado del bosque hacen que se disminuya la velocidad del viento y turbulencia, lo cual ofrece más refugio para el sotobosque creando un ambiente favorable para el establecimiento de la regeneración natural.
En paralelo a otro tipo de bordes como cultivos agrícolas o pastizales, se crean modelos de presión del viento más heterogéneos, sin desaceleración y deflexión del flujo del viento (Agster et al., 2003). Esto genera disminuciones detectables de los valores de humedad y aumento en la temperatura, a los cuales son tolerantes pocas especies generalistas (Meza Elizalde y Armenteras Pascual, 2018) coincidiendo con lo encontrado en este estudio, en bosques con cobertura adyacente a pastos.
Correspondencia entre los ambientes contrastantes
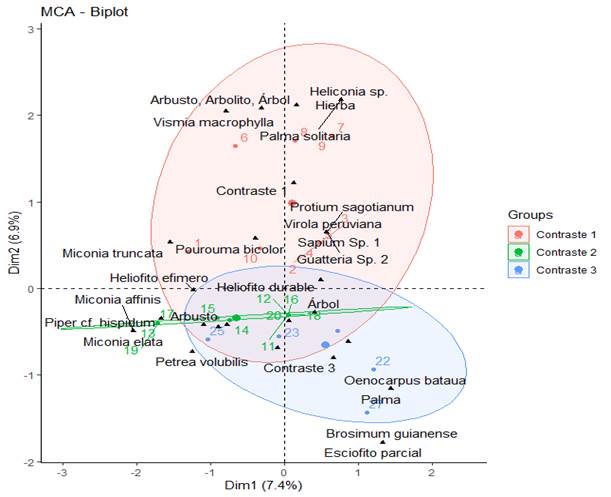
Figura 5. Análisis de correspondencia múltiple para la regeneración natural según el ambiente contrastante, habito y gremio ecológico.
El primer plano factorial (óvalo superior izquierdo) corresponde al ambiente de bosque natural adyacente a cultivo de caucho donde se relacionan las especies de mayor %RG tales como Brosimum guianense, Guatteria cf. metensis, Himatanthus articulatus, Inga acreana, Inga brachyrhachis, Inga. cf. heterophylla, Neea floribunda, Oenocarpus bataua, Otoba Sp. 1, Pausandra trianae, Pseuldomedia laevis, Sorocea steinbachii, Sterculia apetala y Triplaris americana los cuales en su gran mayoría son asociados al habito arbóreo a excepción de la palma Oenocarpus bataua de gremio ecológico heliofito durable con la particularidad de Brosimum guianense que es una especie esciófita parcial, ver Figura 5.
El segundo plano (óvalo al lado derecho del plano) atañe similitud en el ambiente contrastante de bosques con entrada de ganado a las especies Petrea volubilis, Himatanthus articulatus, Miconia elata, Siparuna mutisii y Miconia truncata por su alto %RG, asociadas al habito arbustivo excepto a Himatanthus articulatus y al gremio heliofito efímero, exceptuando de este la especie Siparuna mutisiii la cual es de gremio heliofito durable, ver Figura 5.
El ultimo plano (óvalo en la parte inferior del gráfico) corresponde al ambiente contrastante de bosques adyacentes a pastos sin uso, asociado a las especies Virola peruviana, Pouroma bicolor, Guatteria sp 2, Protium sagotianum, Sapium sp 1, Virola macrophylla, quienes pertenecen al habito arbóreo, y a su vez Heliconia sp, que es una herbácea y Astrocaryum chambira que es una Palma; este eje se asocia al habito arbustivo y al gremio heliofito durable a excepción del Pouroma bicolor y Virola peruviana, ver Figura 5.
En los ejes 1 y 2 (AC3 y AC2), se encontró en común la especie Himatanthus articulatus de gremio heliófito efímero y habito arbóreo, mientras que en el eje 1 y 3 comparten especies del género Guatteria y el hábito palma de las especies Oenocarpus bataua y Astrocaryum chambira del gremio heliofito durable. Solo se asocia una especie de habito herbáceo Heliconia sp en el ambiente contrastante 1. El registro de las palmas, y la hierba en los ambientes AC1 y AC3, se debe posiblemente a la existencia de claros al interior de los bosques estudiados.
El análisis de correspondencia múltiple, confirmó que la vegetación cambia principalmente por las características del ambiente contrastante (bióticas y abióticas), aludidas a su cobertura adyacente. Los bosques sin pastoreo poseen una mayor afinidad entre ellos, en relación a la dominancia del gremio heliófitas durable. Por el contrario, el bosque al cual entra el ganado posee condiciones influenciadas por el pastoreo, generando una dominancia de especies herbáceas y lianas características de ambientes perturbados y que pueden impedir el establecimiento de otras especies forestales. Probablemente, el pastoreo afecte heterogeneidad espacial generando cambios en la estructura y composición de la comunidad de la regeneración a pequeña escala (Morici et al., 2003), al mismo tiempo que el pastoreo genera la compactación del suelo, lo cual, sumado a el daño mecánico del ramoneo, reduce la disponibilidad de sitios seguros para el establecimiento de plántulas (Harper, 1977).
En correspondencia, Hernández et al. (2000) habla que la composición florística es diferencialmente menor en áreas con ganado. Al mismo tiempo, sugiere que la especies que no aparecen en las áreas con pastoreo, son potenciales a extinguirse en presencia de los bovinos, lo cual no podemos precisar en nuestro muestreo, por el tamaño de muestra a comparación, pero si se indica que, de las 281 especies totales, 176 especies no están presentes en el AC2.
La presente investigación aporta a una mayor comprensión de la interacción de la actividad ganadera con atributos poco estudiados, como lo es la regeneración natural del bosque, mostrando que el pastoreo genera cambios sustanciales en la composición, estructura y diversidad de los bosques en estadios sucesionales mayores. Por lo cual, es importante direccionar el manejo integral de las áreas adyacentes a los bosques. En este sentido, la implementación de sistemas silvopastoriles, el cerramiento de bosques degradados para impedir el pastoreo y la reducción de bordes abruptos, se constituyen en herramientas de manejo que permiten la coexistencia de la actividad pecuaria y la conservación de las áreas boscosas, las cuales son importantes para el mantenimiento ambiental y socio ecológico de los territorios.
CONCLUSIONES
Teniendo en cuenta los atributos estructurales florísticos, estructurales y de diversidad de la regeneración natural, se encontró que los bosques sin pastoreo poseen mayor diversidad en la regeneración natural y presencia de especies esciofitas en la dinámica sucesional. En comparación a los bosques con pastoreo, en los cuales se observó un patrón ecológico distintivo de ambientes perturbados, con la presencia de especies generalistas, heliófitas, de hábitos herbáceos y de rápida colonización, lo cual pueden dificultar el reclutamiento de especies forestales. Los bosques adyacentes a plantaciones de caucho, registraron una mayor diversidad de especies forestales de gremios heliófilos y esciofitos. Los atributos ecológicos analizados, indican que probablemente, el pastoreo puede llegar a afectar la heterogeneidad espacial y ecológica de la regeneración natural, creando cambios en la estructura y composición de la comunidad de la regeneración a pequeña escala.
Se sugiere que, en posteriores estudios, se realice la comparación de la abundancia de la regeneración natural con la composición de árboles del estrato superior, para así determinar si el bosque tiene el potencial de mantener su actual composición en el futuro. Así mismo, se recomienda que se hagan análisis del impacto de pastoreo considerando cómo las variables la densidad de ganado que entra al bosque o variables de intensidad de pastoreo, y así articularlo con las comunidades que viven de las actividades agropecuarias y sensibilizarlos con el manejo del recurso bosque y sus servicios ecosistémicos.
Por último, dada la importancia de la regeneración natural en la rehabilitación y restauración de ecosistemas boscosos perturbados o degradados, se sugiere tiempos de monitoreo no inferiores a 10 años.

