











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink
INTRODUCCIÓN
Los bosques constituyen un sistema natural complejo de vital importancia, ya que son proveedores de servicios ambientales indispensables para el mantenimiento de la biodiversidad de los ecosistemas y de regulación del clima a escala regional y global, y constituyen además una fuente de recursos económicos (MEA, 2005; IPCC, 2007; Balvanera, 2012). En Argentina, en el período 2007-2018, se perdieron más de 3,5 millones de hectáreas de bosques nativos, representando la región chaqueña el 80% de la deforestación de 2019 (Unidad de Manejo del Sistema de Evaluación Forestal de la Dirección Nacional de Bosques [UMSEF], 2019). En diciembre de 2018, se renovó la ley 25.080/1999 de Inversiones para Bosques Cultivados (a través de la Ley Nacional 27.487/2018) que, complementada por la Ley Nacional 26.331/2007 de Presupuestos Mínimos de Protección Ambiental de los Bosques Nativos, continúan incentivando la conservación, enriquecimiento, restauración, aprovechamiento y manejo sostenible de los bosques nativos y de los servicios ambientales que estos brindan a la sociedad (Campos et al., 2014). A nivel nacional se cuenta actualmente con 30.359.404 ha de tierras forestales que incluyen bosques y selvas de clima templado y cálido, de las cuales 1.203.674 ha corresponden a bosques cultivados (INDEC, 2018).
La ecorregión del Chaco Seco, caracterizada por sus bosques xerófilos, es considerada como una región forestal, pero debido a la tala indiscriminada, la expansión de la frontera agrícola, el sobrepastoreo, los incendios, la extracción de minerales y petróleo y la urbanización no planificada, estos relictos son intensamente perturbados (Villagra et al., 2009).
Actualmente, la promoción de la implantación de bosques ha ayudado en cierta medida a reducir la tasa de deforestación en la región. Sin embargo, los bosques cultivados constituyen una modificación en el ecosistema, ya que la diversidad de especies vegetales es reemplazada por una plantación monoespecífica, aunque es esperable que estos efectos sean menores si se utilizan especies forestales nativas de la región. Por lo tanto, podría esperarse que la forestación con Prosopis alba Grisebach en la ecorregión del Chaco Seco provoque una menor alteración del ambiente en comparación a la producción de especies forestales exóticas. Una manera de evaluarlo es a través de la comparación de la biodiversidad encontrada en ambos rodales y el empleo de grupos indicadores de artrópodos.
Dado que muchos artrópodos permiten detectar cambios en el funcionamiento de ecosistemas forestales a través del cambio en su distribución, abundancia y composición, el objetivo de este trabajo fue comparar el efecto de bosques implantados de Prosopis alba y Eucalyptus tereticornis Smith sobre la diversidad de artrópodos epígeos en la ecoregión del Chaco Seco.
MATERIALES Y MÉTODOS
Sitio de estudio. El estudio se realizó en la Estación Experimental Fernández, unidad dependiente de la Universidad Católica de Santiago del Estero en el marco del convenio con la provincia de Santiago del Estero, ubicada en la ecorregión del Chaco Seco, Argentina (27,56° S y 63,52° O). La temperatura media es de 20 °C y las precipitaciones anuales promedian los 600 mm con una marcada estacionalidad hacia el verano. Dentro de la estación se seleccionaron rodales experimentales de E. tereticornis y de P. alba de entre 17 y 24 años de edad, y un bosque sin intervenir durante 50 años, compuesto principalmente por Ceiba speciosa A. St.- Hil y Phoenix canariensis Chabaud, seleccionado como referencia debido a la ausencia de bosque nativo en las inmediaciones.
Muestreo. Los muestreos se llevaron a cabo en dos fechas, en el otoño y la primavera del 2019, en coincidencia con las épocas de entrada y salida del reposo invernal de las especies forestales estudiadas. Los insectos se recolectaron mediante trampas de caída, las cuales consistieron en recipientes de plástico de 100 ml, en las que se adicionaron aproximadamente 50 ml de solución de agua con unas gotas de detergente. En el interior de cada rodal se trazaron dos transectas distanciadas entre ellas 10 m, dentro de las cuales se colocaron bajo la copa de seis árboles trampas de caída, con un distanciamiento de 6 m. Por cada tipo de bosque se hicieron tres repeticiones, totalizando 36 trampas por cada uno y 108 en total por estación.
Las trampas permanecieron en su sitio 24 h desde su colocación, luego fueron retiradas y se filtraron los ejemplares capturados. Los artrópodos colectados fueron clasificados como morfoespecies, por sus características en común, dentro de los niveles de Orden y Familia empleando claves dicotómicas para su identificación (Borror y De Long, 1988; Klimaszewski y Watt, 1997; Fernández y Sharkey, 2006; Buck et al., 2009). El material se encuentra preservado en alcohol 70° y almacenado en el Laboratorio de Zoología Agrícola de la Universidad Nacional de Luján.
Análisis de los datos. Se analizaron la abundancia (número de individuos de cada morfoespecie), abundancia relativa (proporción de la abundancia de cada Orden con respecto al número total de individuos), riqueza (número de taxones diferentes), riqueza relativa (proporción de la riqueza de cada Orden con respecto a la riqueza total), equitatividad por el índice de Simpson (λ) y el índice de diversidad de Shannon-Wiener (H’) por cada tipo de bosque. Los índices se obtuvieron de la siguiente manera:
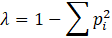
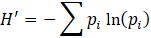
Donde p i es la abundancia relativa de cada taxón.
Para determinar si había un efecto del tipo de bosque sobre la comunidad de artrópodos se empleó la prueba no paramétrica de Kruskal-Wallis (p<0,05). Además, se construyeron curvas de acumulación de morfoespecies a fin de estimar el esfuerzo de muestreo necesario para registrar el mayor número de morfoespecies posibles en cada ambiente. Por otro lado, para determinar la similitud entre sitios se empleó el índice de Jaccard que relaciona el número de morfoespecies compartidos por cada comunidad con el total, por lo tanto, valores cercanos a 1 indican mayor similitud.
Todos los análisis fueron realizados por medio del software RStudio (R Core Team, 2019).
RESULTADOS Y DISCUSIÓN
Se recolectó un total de 2.291 individuos distribuidos en 72 morfoespecies, 912 individuos de 44 morfoespecies durante el otoño y 1.379 individuos de 53 morfoespecies en primavera (Tabla 1). Al observar las curvas de acumulación de morfoespecies (Figura 1), se aprecia que para los sitios de E. tereticornis y monte sin intervención, el número de unidades muestrales empleado parece ser el adecuado ya que las curvas reducen notablemente su pendiente. Cabe aclarar que la curva del monte sin intervención es de menor longitud porque no se pudieron recuperar las trampas en su totalidad. Sin embargo, para P. alba sería conveniente la inclusión de más unidades de muestreo ya que la pendiente se mantiene constante, lo que indica que aún quedan morfoespecies por identificar.
Al analizar la abundancia, la riqueza y los índices, se encontró que durante la primavera los valores fueron mayores en P. alba con respecto a E. tereticornis y al monte sin intervención, mientras que durante el otoño no se observaron diferencias significativas, indicando que las comunidades asociadas a cada sitio en esta estación podrían tener una estructura similar (Tabla 2). Por su parte, la mayor diversidad de artrópodos terrestres asociada al bosque de P. alba observada en primavera coincide con lo encontrado por Penon (2018), quien señala que los montes de eucalipto poseen una menor diversidad de artrópodos con respecto a Robinia pseudoacacia L., una leguminosa arbórea fijadora de nitrógeno que produce residuos de mayor valor nutritivo, favoreciendo a la proliferación de otras especies vegetales que podrían servir como recurso para algunos grupos de insectos.
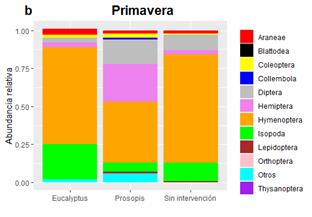
En ambas estaciones, la riqueza de morfoespecies estuvo distribuida de forma más homogénea entre los órdenes (Figura 2) que la abundancia a nivel de orden (Figura 3), lo que se tradujo en valores bajos de equitatividad (Tabla 2). En los tres sitios y en ambas épocas dominó el orden Hymenoptera (Figura 3), en una proporción entre 40 y 71 %, siendo la familia más abundante Formicidae, lo cual coincide con valores encontrados por otros autores para bosques tropicales y subtropicales de ambientes secos (Forbes, Van Zee, Smith y Whitford, 2005; Diodato & Fuster, 2016). Esta dominancia tan marcada se podría deber a la oferta de recursos alimenticios que ofrecen las plantas en primavera, que favorecen la búsqueda de alimento por parte de las hormigas (Jaffé et al., 2007; Diodato & Fuster, 2016). Por otro lado, los ecosistemas de bosques benefician la actividad de la macrofauna e ingenieros del suelo, entre los que se incluyen las hormigas, debido a la mayor disponibilidad de carbono orgánico, la mayor cantidad de hojarasca depositada y la relativamente baja perturbación de este tipo de uso del suelo con respecto a actividades agrícolas y ganaderas (Penon, 2018). En el rodal de P. alba, Hemiptera aumentó su abundancia de 2 a 25 % entre fechas, siendo las familias Psyllidae y Cicadellidae las más abundantes. El predominio de estas dos familias se debe a la estrecha relación existente entre sus abundancias y la estacionalidad de los recursos, ya que, por su régimen alimentario, estas especies requieren de tejidos vegetales con un flujo activo de savia. El mismo es mayor en primavera, cuando las plantas están en pleno crecimiento, lo cual promueve a que el Orden Hemiptera constituya uno de los principales grupos de herbívoros que se sustentan de estos bosques (Neves, Sperber, Campos, Soares & Ribeiro, 2013; Riquelme Virgala et al., 2016). En los bosques de E. tereticornis y sin intervención, fue más dominante el orden Isopoda, probablemente debido a que la menor tasa de descomposición de la hojarasca genera un ambiente propicio para el desarrollo de estos organismos detritívoros (Sayer, Sutcliffe, Ross & Tanner, 2010; Diodato & Fuster, 2016; Penon, 2018).
Los resultados de este trabajo indicarían que la forestación con P. alba en la ecorregión del Chaco Seco provoca una menor alteración del ambiente en comparación a la producción de especies forestales exóticas. Sin embargo, la diversidad de artrópodos asociada parece ser menor que la del bosque nativo. En un estudio realizado por Diodato y Fuster (2016) en diferentes localidades de la ecorregión Chaco Seco, en el que se evaluó la diversidad de artrópodos del dosel del bosque nativo, registraron una riqueza de insectos superior a la observada en este trabajo para el monte de P. alba, aunque con una participación similar de algunos grupos como Hymenoptera y Hemiptera. Por su parte, Baudino, Cecchetto, Buffa y Visintin (2020) registraron la diversidad de fauna epígea asociada a distintos ambientes de la misma ecorregión y observaron que el bosque nativo presentó la mayor abundancia y riqueza de insectos debido a la heterogeneidad vegetal, también alcanzando valores superiores a los vistos para el rodal de P. alba evaluado en este trabajo.
La baja diversidad observada en el bosque tomado como referencia podría ser un indicador de que las especies vegetales que lo componen no proveen características de hábitat similares al bosque nativo. Esto se ve reflejado en la similitud de sus valores de riqueza, abundancia y diversidad con respecto al rodal de E. tereticornis. Por lo tanto, a pesar de ser un sitio sin intervención humana durante muchos años, este ambiente no puede ser tenido en cuenta como bosque de referencia representativo de la artropodofauna local. Nuevos relevamientos en los que se incluyan áreas de bosque nativo se consideran necesarios para corroborar los resultados de este trabajo, que indican que el bosque cultivado de P. alba favorece la conservación de la biodiversidad en la ecorregión Chaco Seco en comparación con montes forestales de especies exóticas.
Tabla 1 Listado de morfoespecies encontradas con sus respectivas abundancias, en tres tipos de bosque en la ecorregión Chaco Seco durante dos estaciones.
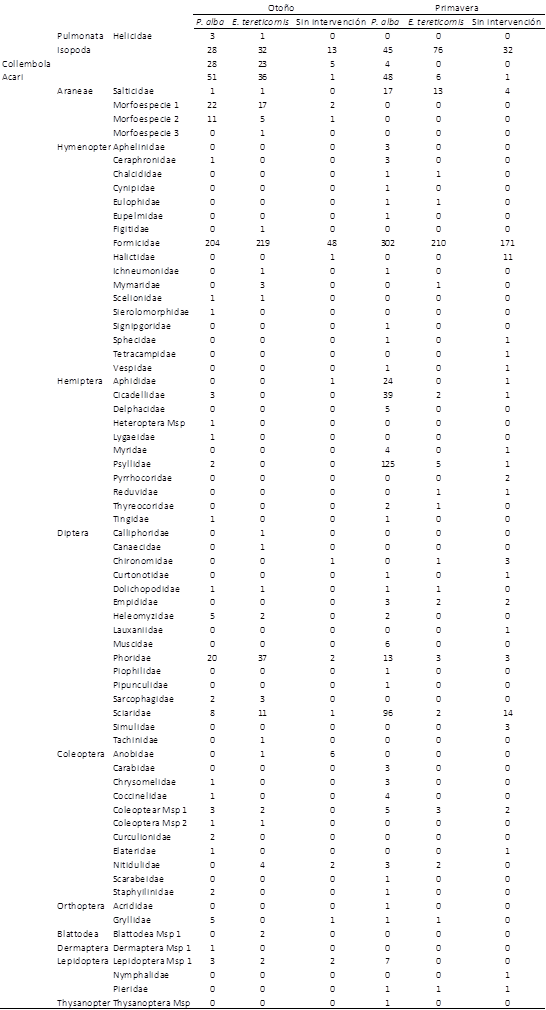
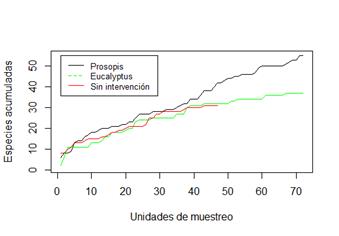
Figura 1 Curvas de acumulación de morfoespecies encontradas en cada sitio durante todo el período de muestreo.
Tabla 2 Valores de riqueza, abundancia e índices de Simpson y Shannon-Wiener estacional en tres tipos de bosque en la ecorregión Chaco Seco. Letras diferentes en una misma columna indican diferencias estadísticamente significativas (LSD de Fisher, p <0.05).
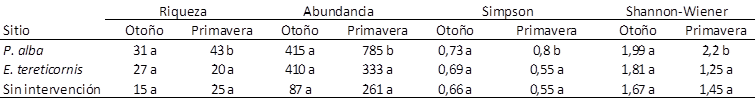
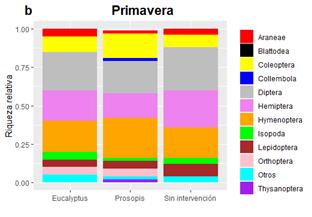
Figura 2 Riqueza relativa de morfoespecies de los distintos órdenes de Arthropoda encontrados en cada sitio durante (a) otoño y (b) primavera.
CONCLUSIONES
A través de este estudio preliminar se observa que el bosque cultivado de P. alba parece favorecer la conservación de la biodiversidad de artrópodos epígeos en la ecorregión del Chaco Seco, Argentina. Nuevos estudios se consideran necesarios para afianzar estos resultados y profundizar en la caracterización de la diversidad asociada a estos bosques, así como también desarrollar modelos que permitan evaluar el estado de conservación de estos ecosistemas a través de indicadores biológicos.

