











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
El cultivo de trigo en Paraguay ha logrado grandes avances en su producción durante la última década. Sin embargo, dicho logro está amenazado anualmente por diversas limitaciones incluyendo las epidemias de enfermedades. Una de estas, conocida como pyricularia o brusone causado por Magnaporthe oryzae B. C. Couch and L. M. Kohn (sin. Pyricularia oryzae). Este hongo, presenta una amplia variabilidad patogénica (Zeigler et al. 1995), existen varias cepas, las cuales tienden a variar el grado de especificidad hospedera, y pueden ser divididas en patotipos en base a la preferencia de hospederos. Las cepas que comúnmente atacan al trigo en Sudamérica, han sido denominadas como Magnaporthe oryzae Triticum pathotype (MoT) (Valent et al. 2016, Cruz et al. 2016).
Desde su primer reporte a nivel mundial en Brasil en 1985 (Igarashi et al. 1986), esta enfermedad ha causado varias epidemias en Brasil, Bolivia y Paraguay, siendo considerada la mayor amenaza a la producción de trigo en Sudamérica (Kohli et al. 2011). Recientemente la enfermedad ha sido reportada en Bangladesh causando epidemias devastadoras (Malaker et al. 2016) y es considerada como una amenaza potencial a la producción de trigo en los Estados Unidos (Cruz et al. 2016). Todo esto la convierte en una de las más importantes enfermedades del trigo en la actualidad.
En Paraguay, la primera epidemia fue observada en el año 2002, causando severas pérdidas a la producción nacional (Viedma y Morel 2002). Según Viedma (2010), las siembras tempranas (primera quincena de abril), tienen mayor posibilidad de ataque si se producen condiciones de alta temperatura y humedad en los meses de junio y julio. Mientras que en años muy húmedos con prevalencia del fenómeno El Niño, los ataques de Pyricularia pueden ocurrir independientemente de la fecha de siembra (Kohli et al. 2011).
Actualmente son pocos los materiales que han sido reportados con altos niveles de resistencia: BR 18, Sausal CIAT, Paragua CIAT, Milan, CD 116 (Kohli et al. 2011, Fronza et al. 2016). Dichos materiales fuentes forman parte del programa nacional de mejoramiento. Sin embargo, poco se conoce acerca del comportamiento de los trigos nacionales y de las líneas derivadas del cruzamiento de las fuentes de resistencia mencionadas.
Numerosas investigaciones, han sido realizadas, para identificar fuentes de resistencia y desarrollar estrategias de control que permitan reducir las pérdidas ocasionadas por la enfermedad (Urashima et al. 2004, Arruda et al. 2005, Prestes et al. 2007). La mayoría de estas investigaciones fueron basadas en la infección foliar (Urashima y Kato 1994; Metha et al. 2001; Urashima et al. 2004; Marangoni et al. 2013). Considerando que la enfermedad es más severa en el estadio reproductivo afectando a la espiga y los granos, este trabajo buscó relacionar la resistencia a la infección en las hojas con la resistencia en las espigas, ya que se observan varias diferencias en la expresión de la resistencia a nivel de campo. Dependiendo de la validez de la infección foliar, se podría evaluar mayor cantidad de materiales en menor tiempo y ante una mayor cantidad de aislados del hongo. Esto permitiría ampliar la base de resistencia a Pyricularia en corto a mediano plazo.
MATERIALES Y MÉTODOS
El ensayo se realizó en las instalaciones del Centro de Investigación Hernando Bertoni del Instituto Paraguayo de Tecnología Agraria (IPTA), durante el año 2014. Sesenta y un líneas avanzadas y variedades de trigo, representando una gran diversidad genética del Programa de Investigación de Trigo del IPTA, fueron evaluadas bajo condiciones semi controladas (Tabla 1).
El ensayo utilizó un diseño completamente al azar con cuatro repeticiones, tanto en la infección foliar como en las espigas, considerando una maceta con cuatro plantas como unidad experimental.
La inoculación foliar se llevó a cabo en el estado de cuatro hojas, y la de espigas cuando las mismas se encontraban completamente fuera de la hoja bandera.
El aislado de Pyricularia oryzae empleado para las inoculaciones fue P13-009, proveniente de las colecciones de origen Capitán Miranda, Itapúa, durante el ciclo 2013. Este aislado fue conservado sobre papel de filtro estéril a -18°C (Aricapa y Correa 1994), repicado a placas de Petri con medio de cultivo de avena e incubado durante 12 días a 25°C con fotoperiodo de 12 horas. Posteriormente el micelio fue aplastado con un triángulo de vidrio y las placas incubadas con luz constante por 72 horas para favorecer la esporulación (Marangoni et al. 2013). El conteo de conidios se realizó empleando un hematocitómetro, y la concentración fue ajustada a 2 105 conidios mL-1.
Las inoculaciones fueron realizadas asperjando las hojas y espigas con un aspersor común. Posterior a la aspersión las plantas fueron mantenidas en oscuridad por 24 horas, en una sala climatizada con 80% de humedad y 28°C de temperatura. Posterior a esta incubación las plantas fueron trasladadas a un invernadero con temperatura de 26 ± 3°C.
Tabla 1 Variedades y líneas de trigo utilizadas en el ensayo y su número de referencia. Caacupé, 2014.


Figura 1 Escala utilizada para la evaluación de las infecciones causadas por P. oryzae en hojas de trigo adaptada de Valent et al. (1991) (Caacupé, 2014).
Infección foliar: la evaluación foliar se realizó en la cuarta hoja de cada planta ocho días después de la inoculación. Para la evaluación de las lesiones foliares causadas por P. oryzae en plantas de trigo, fue adaptada la escala propuesta por Valent et al. (1991) (Figura 1), de la siguiente manera para la toma de notas:
0 = Sin infección
1 = Lesiones puntuales del tamaño de una cabeza da alfiler de color marrón
2= Lesiones elípticas o estriadas de 2 mm o más sin centro distinguible
3 = Lesiones redondeadas con borde oscuro y centro ceniza, en algunos casos con halo amarillo
4 = Lesiones típicas del brusone, elípticas con borde oscuro y centro ceniza
5= Grandes lesiones elípticas con centro gris oscuro
Las lesiones con escala 0, 1 y 2 son consideradas de tipo resistente, debido a la ausencia de esporulación en las condiciones favorables y las lesiones 3, 4 y 5 son consideradas de tipo susceptible (Valent et al. 1991; Urashima et al. 2004).
En base a la frecuencia del tipo de lesiones observadas en cada material, los mismos fueron clasificados de la siguiente manera:
Resistente (R): 100% de las lesiones de tipo 0, 1 y 2
Moderadamente resistente (MR): Más del 50% de las lesiones de tipo 0, 1 y 2
Moderadamente susceptible (MS): Más del 50% de las lesiones de tipo 3, 4 y 5
Susceptible (S): 100% de las lesiones de tipo 3, 4 y 5
Infección en espigas: La evaluación se realizó 15 días después de la inoculación, para ello fue elaborada una escala considerando el porcentaje de la espiga necrosada, la misma se presenta en la Figura 2, y se clasificó considerando la siguiente escala:
0 = Sin infección
1 = Hasta el 10% de la espiga necrosada
2 = Hasta el 40% de la espiga necrosada
3 = Hasta el 60% de la espiga necrosada
4 = 100% de la espiga necrosada

Figura 2 Escala de severidad utilizada para la evaluación de la infección causada por Pyricularia en materiales de trigo (Caacupé, 2014).
Teniendo en cuenta la nota máxima alcanzada por cada material, los mismos se clasificaron en: resistentes, cuando la nota máxima fue 1; moderadamente resistentes cuando la nota máxima fue 2; moderadamente susceptibles cuando la nota máxima fue 3 y susceptibles cuando la nota máxima fue 4. Los datos de ambas evaluaciones fueron sometidos al análisis de varianza y la comparación de medias por la prueba de LSD de Fisher α = 0,05 utilizando el programa estadístico Infostat (Di Rienzo et al. 2015). El análisis de correlación de Pearson fue utilizado para establecer si existe una relación entre el comportamiento de los materiales en los dos estadios evaluados.
RESULTADOS Y DISCUSIÓN
Los resultados de las inoculaciones controladas en el invernadero mostraron diferencias estadísticas significativas entre los diferentes materiales genéticos, sometidos a la inoculación foliar y en espigas. Para la infección foliar, 35 de los materiales fueron clasificados como resistentes, 8 moderadamente resistentes, 10 moderadamente susceptibles y 8 susceptibles. Sin embargo, la evaluación de la infección en estado de espigzon resulto ser muy diferente a aquella observada en la hoja. En este caso, solo cinco de los materiales genéticos fueron clasificados como resistentes, cuatro moderadamente resistentes, 13 moderadamente susceptibles y 39 susceptibles. Los datos presentados en las Tablas 2, 3, 4 y 5 muestran la clasificación de los materiales basados en ambas inoculaciones y su reacción a la inoculación foliar y en espiga.
Tabla 2 Variedades y líneas de trigo clasificados como resistentes a la infección foliar y su reacción a la infección en espigas. Caacupé, 2014.

*Medias con una letra común no son significativamente diferentes (p > 0,05). Test:LSD Fisher Alfa=0,05; X1: Clasificación según tipo de lesión en hoja; X2: Clasificación según severidad en espiga.
De los 35 materiales clasificados como resistentes a la infección foliar, solo cinco resultaron ser resistentes y uno moderadamente resistente en el estadio de espiga (Tabla 2). Ninguno de los materiales tipificados como moderadamente resistentes en la infección foliar mantuvo su resistencia en la espiga (Tabla 3). De todos los materiales clasificados como moderadamente susceptibles o susceptibles a la infección foliar, solo dos, uno en cada categoría, fueron clasificados como moderadamente resistentes a la infección en espiga (Tablas 4 y 5).
Considerando ambas infecciones, los materiales que presentaron resistencia tanto a nivel foliar como en la espiga fueron los genotipos de referencia N° 9, 14, 30, 45 y 59. De entre estos materiales, sólo la línea N° 59 (Milan) es mencionada como fuente de resistencia por Kohli et al. (2011). Estos autores también señalan a Paragua CIAT y Parapeti CIAT como resistentes, y BR 18, IAPAR 85 y CD 113 como moderadamente resistentes. Sin embargo, el presente estudio BR 18 y Parapeti CIAT fueron clasificados como resistentes a nivel foliar, pero susceptibles y moderadamente susceptibles en las espigas.
Tabla 3 Variedades y líneas de trigo clasificados como moderadamente resistentes a la infección foliar y su reacción a la infección en espigas. Caacupé, 2014.
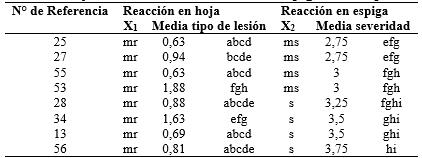
*Medias con una letra común no son significativamente diferentes (p > 0,05). Test:LSD Fisher Alfa=0,05; X1: Clasificación según Tipo de lesión en hoja; X2: Clasificación según severidad en espiga.
Tabla 4 Variedades y líneas de trigo clasificados como moderadamente susceptibles a la infección foliar y su reacción a la infección en espigas. Caacupé, 2014.
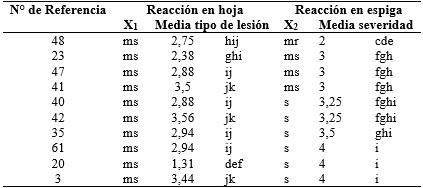
*Medias con una letra común no son significativamente diferentes (p > 0,05). Test:LSD Fisher Alfa=0,05; X1: Clasificación según Tipo de lesión en hoja; X2: Clasificación según severidad en espiga
Tabla 5 Variedades y líneas de trigo clasificados como susceptibles a la infección foliar y su reacción a la infección en espigas. Caacupé, 2014.

*Medias con una letra común no son significativamente diferentes (p > 0,05). Test:LSD Fisher Alfa=0,05; X1: Clasificación según Tipo de lesión en hoja; X2: Clasificación según severidad en espiga.
Arruda et al. (2005) observaron resultados similares en BR 18, la cual, a pesar de ser su susceptibilidad en Paraguay, sigue siendo considerada como línea de moderada a alta resistencia por los investigadores en Brasil (Prestes et al. 2007, Storani 2013, Fronza et al. 2016). Entre las líneas avanzadas descendientes de Paragua CIAT y Milan, desarrolladas por el Programa Nacional de Trigo en Paraguay (referencia 49, 50 y 51), solo la N° 50 (PARAGUA CIAT//MILAN/MUNIA), presentó resistencia a la inoculación foliar y moderada resistencia en espigas. Este hecho puede indicar la complejidad en el manejo de las fuentes de resistencia genética solo en base a las observaciones del campo y la dificultad para combinarlas con el objetivo de mejorar los niveles de resistencia.
La frecuencia de reacción a la infección foliar y en espiga de los genotipos evaluados en este estudio, es presentada en la Tabla 6. Los resultados demuestran que, de los 35 materiales resistentes a la infección foliar, 28 mostraron susceptibilidad en las espigas, y solo siete de los 61 materiales estudiados se comportaron entre moderadamente resistentes y resistentes a ambas infecciones.
Tabla 6 Frecuencia de reacción de los 61 genotipos de trigo a la infección foliar y en espiga. Caacupé, 2014.
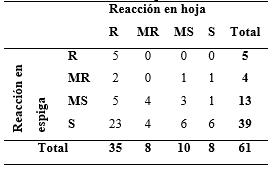
Estos datos confirman la falta de relación entre las infecciones foliares y en espiga (Pearson Coef.: 0,10618/p=0,61346). Las observaciones de Arruda et al. (2005) que clasifican solo 3 de los 15 materiales resistentes en estadio foliar a susceptible en espiga es un bajo indicativo del problema en evaluación del germoplasma en hojas. En este estudio, de los 35 materiales considerados resistentes en la hoja, la mayoría (28) fue encontrada susceptible o moderadamente susceptible en estado de la espiga. Esta contundencia de resultados confirma, en general, la falta de relación entre las reacciones de Pyricularia en la hoja con aquellos en la espiga.
Estos resultados realzan las observaciones empíricas de campo, donde identificar materiales resistentes a Pyricularia en estado de espiga ha sido una tarea difícil hasta ahora. Considerando la alta coincidencia de la susceptibilidad en la hoja con susceptibilidad en la espiga, la reacción en la hoja puede servir para descartar los materiales susceptibles y así, limitar los estudios de la espiga solo sobre aquellos materiales con reacciones MS, MR y R en la hoja. Esto permitiría evaluar materiales con mayor probabilidad de presentar resistencia en espigas e introducir nuevas fuentes de resistencia en los programas de mejoramiento.
En base al bajo número de líneas consideradas resistentes a Pyricularia en este estudio, se enfatiza la necesidad de ampliar resistencia genética en el germoplasma de trigo de manera urgente.
Finalmente cabe poner énfasis que los datos reportados aquí provienen de las inoculaciones con un solo aislado del hongo, considerado virulento en condiciones controladas y de campo. Es totalmente posible que los materiales identificados como resistentes en este estudio, tengan una reacción diferenciada con otros aislados del patógeno y en el campo, dependiendo del complejo patogénico y las condiciones ambientales de la localidad.
CONCLUSIÓN
Este estudio encuentra que la resistencia foliar al brusone no es representativo de la resistencia en las espigas. Sin embargo, la susceptibilidad en la hoja, en general, es indicador de la susceptibilidad en la espiga. Los resultados logrados en base a un gran número de materiales genéticos utilizados por el Programa Nacional de Mejoramiento de Trigo en Paraguay demuestran la estrechez del germoplasma resistente a las infecciones de Brusone en la espiga que representa una limitación seria. Se sugiere ampliar la búsqueda de germoplasma resistente y su incorporación en el programa de manera urgente.

