











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
Si bien habitualmente el fenotipo se expresa solo en términos de genotipo y medio ambiente bajo el supuesto que dichos componentes actúan en forma independiente, en ocasiones ello no se cumple debido a la presencia de correlación y/o interacción entre los mismos. La interacción mencionada puede ser descripta como la respuesta diferencial de los genotipos ante un cambio de ambiente - entendiéndose a este último en un sentido amplio - lo que agregaría una fuente de variación adicional a la variancia fenotípica total 1,2. En trabajos pioneros 3 se observó que es de esperar que el significado de la interacción sea mayor cuanto más difieren entre sí los genotipos involucrados y los ambientes considerados. En este sentido, diferentes autores documentaron un efecto significativo de la interacción genotipo, nivel nutricional y localidad sobre el crecimiento tanto en aves de carne 4 como de postura 5. En tal sentido, se ha informado que la exposición de parrilleros comerciales a temperaturas por encima de la zona de termoneutralidad afecta la respuesta fisiológica y el desempeño productivo 6,7 y modifica el contenido proteico de la pechuga de las aves 8.
Esta evidencia ha llevado a considerar la introgresión de genes mayores vinculados con la termorregulación como estrategia genética para contrarrestar dicho impacto 9,10. En estudios de ecotipos autóctonos sometidos a dos sistemas de crianza contrastantes en comunidades rurales de Tanzania 11 se observó interacción genotipo x manejo significativa en relación al peso corporal de las aves en diferentes edades en contraposición a lo informado por otros autores 12 en investigaciones similares. El pollo campero es un recurso genético apto para sistemas avícolas alternativos 13 y en Argentina es distribuido por el Programa Pro-Huerta para la autoproducción de alimentos y la venta de excedentes por parte de poblaciones nutricionalmente vulnerables. El mismo representa, además, una alternativa de diversificación para productores a pequeña escala, destinado a consumidores que demandan alimentos generados en condiciones más naturales que prioricen aspectos de la etología y el bienestar animal. A nivel institucional, el INTA (Instituto Nacional de Tecnología Agropecuaria) tiene a su cargo la distribución de pollos camperos de genética propia a diferentes zonas del país y si bien este tipo de ave constituye un recurso genético con mayor variabilidad adaptado a condiciones de cría más diversas, en esta situación cobran particular trascendencia las posibles interacciones genotipo x localización geográfica 14.
El objetivo de este estudio fue evaluar el efecto de dos estaciones contrastantes del año, sobre el comportamiento productivo de machos de pollo campero derivados de dos modelos genéticos de cruzamientos terminales: Campero INTA Tradicional, producto de un cruzamiento simple y Campero Casilda, producto de un cruzamiento experimental de tres vías.
MATERIAL Y MÉTODOS
Las aves fueron producidas en la Estación Experimental Agropecuaria de INTA (EEA “Ing. Agr. Walter Kugler”) ubicada en la localidad de Pergamino, Provincia de Buenos Aires, Argentina, en dos estaciones de crianza: otoño-invierno (abril a julio) y primavera (setiembre a diciembre) con diferente impacto de las variables medioambientales (temperaturas media, máxima y mínima y humedad relativa - otoño: 13,5°C, 16,6°C, 9,2°C, 82 %; primavera: 19,4°C, 22,5°C, 16,4°C y 72,5 %) En cada época, se utilizaron cuarenta pollos machos de cada uno de los siguientes grupos genéticos: Campero Casilda (CC) y Campero INTA (CI). Campero Casilda es un cruzamiento experimental de tres vías entre gallos de la población sintética paterna AH' y gallinas derivadas del cruzamiento simple entre gallos y gallinas de las poblaciones sintéticas maternas A y ES, respectivamente. Campero INTA es un cruzamiento simple entre gallos de la población sintética paterna AS y gallinas de la población sintética materna E. Las poblaciones intervinientes se generaron y mantienen en el Núcleo Genético de la Sección Avicultura de la EEA Pergamino del INTA. La constitución genética teórica de las mismas (Bonino, comunicación personal) es: Sintética materna A - 75% Cornish Colorado + 25% Rhode Island Red; Sintética materna ES - 87,5% Cornish Colorado + 12,5% Rhode Island Red, Sintética materna E - 50% Cornish Colorado + 50% Rhode Island Red; Sintética paterna AH' - población mejorada por velocidad de crecimiento y eficiencia de conversión de alimento a partir de la sintética AH (50% Hubbard + 50% estirpe Anak (grises)) y Sintética paterna AS (50% Cornish Colorado + 50% Cornish Blanco). El día del nacimiento, las aves se sexaron por inspección de la cloaca, se vacunaron contra la enfermedad de Marek, se identificaron con una banda alar numerada y se trasladaron a la Facultad de Ciencias Veterinarias de la Universidad Nacional de Rosario, en la localidad de Casilda (33º 02´ 39´´ de latitud sur, 61º 10´ 05´´ de longitud oeste), donde fueron sometidas al manejo y al plan sanitario establecido en el protocolo de Campero INTA 15. Durante la fase experimental del proyecto, común a las dos épocas del año, 80 aves de cada genotipo se criaron en confinamiento como un único grupo hasta los 35 días de edad con una densidad promedio de 15 aves/m2. A partir del día 36, cuarenta aves elegidas al azar de cada grupo genético, se alojaron en corrales ubicados dentro de un galpón, con acceso a espacios abiertos con una densidad de 2 aves por m2 en la zona de parque, donde se mantuvieron hasta la faena a los 84 días de edad.
Los animales recibieron el mismo manejo de la alimentación, basado en sólo dos tipos de alimento (Iniciador y Terminador) de acuerdo al siguiente detalle: Iniciador (100%) entre el nacimiento y los 28 días de edad; una mezcla de Iniciador (75%) + Terminador (25%) entre los 29 y los 42 días de edad; una mezcla de Iniciador (50%) + Terminador (50%) entre los 43 y los 56 días de edad; una mezcla de Iniciador (25%) + Terminador (75%) entre los 57 y los 70 días de edad y, por último, Terminador (100%) entre los 71 y los 83 días de edad. La formulación de las raciones se llevó a cabo según exigencias del protocolo vigente, con la siguiente composición: Iniciador - proteína 18,5%, calcio 0,9%, fósforo total 0,75%, fósforo disponible 0,47%, energía metabolizable aves 3150 kcal., metionina + cistina 0,72% y lisina 0,94% y Terminador - proteína 15,09%, calcio 0,85%, fósforo total 0,6%, fósforo disponible 0,38%, energía metabolizable aves 3350 kcal., metionina + cistina 0,60% y lisina 0,75%. A intervalos semanales entre el nacimiento y la faena se registró el peso de cada ave con aproximación al gramo (0-35 días) y con aproximación a los 10 gramos (36-84 días). Los datos longitudinales peso corporal-edad se ajustaron por regresión no lineal con el modelo sigmoideo de Gompertz (16: W (t) = A exp (-b exp (- k t)), donde: W (t) = peso corporal (g) en el tiempo t, A = peso corporal asintótico (peso corporal promedio cuando t tiende a infinito), b = parámetro de posición, constante de integración sin significado biológico, que ajusta para aquellos casos en que t es distinto de cero, k = tasa de maduración (velocidad de aproximación al valor A) y t = tiempo en semanas. La bondad del ajuste se evaluó a partir de su convergencia en una solución, del valor del coeficiente de determinación no lineal (R2) ajustado y del comportamiento aleatorio de los residuales determinado mediante un test de rachas o ciclos 17. A los efectos del tratamiento estadístico de los datos, los estimadores de los parámetros A y k se consideraron como nuevas variables aleatorias.
En cada época y a partir de los 36 días de edad, un segundo grupo de 20 aves de cada grupo genético se alojó en jaulas individuales para el control del consumo voluntario de alimento. Luego de un período de acostumbramiento de siete días, cada ave fue pesada semanalmente, se calculó su aumento medio diario de peso (AMD) y se determinó su consumo medio diario individual de alimento (CMD) a partir del registro de la diferencia entre una cantidad fija ofrecida y la cantidad remanente en el comedero al día siguiente. El comportamiento de la relación de conversión (CMD/AMD) y las variables que la determinan se evaluó entre los 42 y los 77 días de edad.
En cada época del año, 20 aves de 84 días de edad, de cada grupo genético, extraídas al azar de las 40 alojadas en galpones con acceso a parque se trasladaron para su faena a la Sección Avicultura de la EEA INTA Pergamino. Se determinó el peso corporal pre-faena, el peso eviscerado, el peso de la pechuga con hueso y de la pata-muslo derecha y el peso del depósito graso abdominal como estimador del contenido de grasa corporal. Los porcentajes de pechuga, pata-muslo y grasa abdominal se calcularon como proporción del peso corporal eviscerado. El rendimiento de la canal (%) se calculó como((peso corporal eviscerado / peso vivo pre-faena) x 100). Los efectos del grupo genético, la época del año y la interacción simple entre ambos factores principales sobre las diferentes variables respuesta, se evaluaron con un análisis de la variancia correspondiente a un diseño completamente aleatorizado con un experimento factorial 2x2 (dos grupos genéticos x dos ambientes climáticos). Se consideraron significativos los efectos con probabilidades asociadas ≤ 0,05.
RESULTADOS Y DISCUSIÓN
La Tabla 1 presenta los valores de los estimadores de los parámetros de la función de Gompertz. Los datos mostraron un comportamiento compatible con el modelo propuesto, el coeficiente de determinación no lineal ajustado (R2) superó el valor de 0,90 y los residuales mostraron una distribución aleatoria (P > 0,05). No se observaron efectos significativos de la interacción simple grupo genético x estación de crianza (F = 0,041; P = 0,841) en relación al peso asintótico de las aves. El efecto del grupo genético tampoco representó una fuente significativa de variación (F = 0,091; P = 0,760) mientras que sí lo fue la estación de crianza (F = 44,3; P < 0,0001) correspondiendo mayores valores del parámetro a las aves criadas en otoño con una depresión del peso asintótico de 13,5 % en Campero Casilda y del 12,7% en Campero INTA ante el cambio de estación y siendo este efecto común a ambos grupos genéticos dada la ausencia de interacción.
Tabla 1. Estimadores de la función de Gompertz aplicada al ajuste de los datos peso corporal-edad cronológica en dos genotipos de pollos camperos.
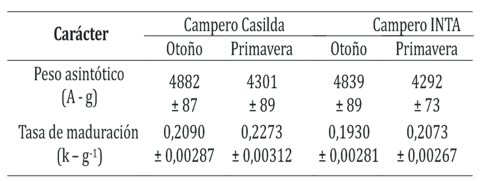
Todos los valores corresponden a la media aritmética ± error estándar
Tamaño muestral: n = 40 aves por subgrupo genotipo x estación
En relación a la tasa de maduración, la ausencia de interacción significativa (F= 0,480; P= 0,487) permitió evaluar los efectos de los factores principales. El grupo genético afectó significativamente (F = 39,3; P < 0,0001) el valor promedio del parámetro que fue mayor en Campero Casilda, independientemente de la estación de año considerada. El efecto estación de crianza también resultó estadísticamente significativo (F= 32,2; P < 0,0001) mostrando mayor tasa de maduración para el carácter las aves de uno y otro grupo criadas en primavera (Campero Casilda: 8,8% y Campero INTA: 7,4%) y siendo dicho efecto de igual magnitud en los dos genotipos considerados.
La Tabla 2 presenta los valores de relación de conversión y de las variables asociadas que determinan su valor. La ausencia de interacción simple estadísticamente significativa en el caso del consumo medio diario de alimento (F= 1,58; P = 0,212) y del aumento medio diario de peso corporal (F= 0,01; P = 0,912) permitió evaluar los efectos directos de los dos factores principales. El CMD fue menor en primavera en ambos grupos genéticos. En otoño, las aves de ambos genotipos mostraron valores similares de consumo, mientras que en primavera las diferencias entre ellos aumentaron, mostrando el genotipo de referencia -Campero INTA- mayor depresión debido al impacto de las condiciones climáticas. Pese a que el comportamiento mencionado en el CMD sugiere la presencia de interacción, la misma no alcanzó significado estadístico. El AMD mostró en ambos genotipos menores valores en primavera sin cambio de comportamiento de los genotipos respecto del desempeño en otoño. La vinculación de ambas variables en la relación de conversión, permitió constatar una interacción pese a que la misma no alcanzó significado estadístico (F=3,06; P =0,084).
Tabla 2. Relación de conversión y variables asociadas de dos genotipos de pollos camperos criados en dos estaciones del año.
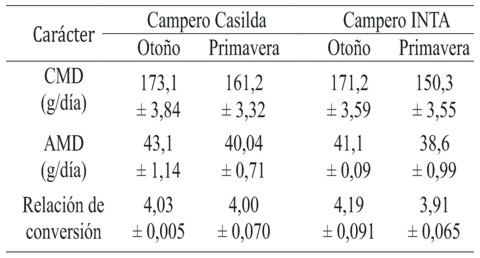
Todos los valores corresponden a la media aritmética ± error estándar
Tamaño muestral: n = 20 aves por subgrupo genotipo - estación del año
La Tabla 3 resume los valores de los caracteres estudiados a la faena. Se constató un efecto significativo de la interacción grupo genético x manejo climático (P < 0,05) en la proporción de pechuga, dado que Campero Casilda presentó mayor proporción de ese corte durante la crianza en primavera a diferencia de Campero INTA genotipo en el cual la mayor proporción se observó en otoño. Para el resto de las variables las aves de los dos genotipos mostraron, en promedio, similar comportamiento independientemente de la estación de crianza.
Tabla 3. Caracteres a la faena de dos genotipos de pollos camperos criados en dos estaciones del año.
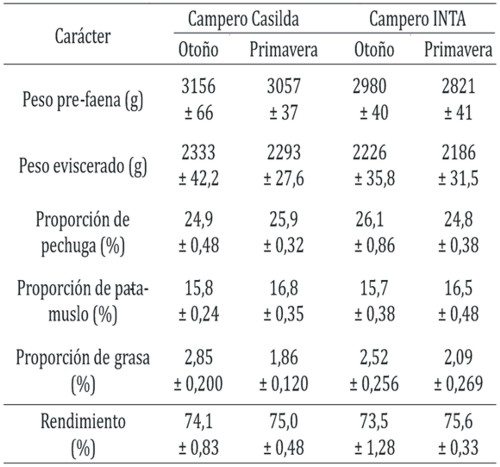
Todos los valores corresponden a la media aritmética ± error estándar
Tamaño muestral: n = 20 aves por subgrupo genotipo - estación del año
El grupo genético representó una fuente de variación significativa para el peso corporal pre-faena (F = 13,10; P < 0,001) y eviscerado (F = 9,5; P = 0,003) correspondiendo mayores valores al cruzamiento experimental. El efecto de la estación del año fue estadísticamente significativo sobre la proporción de pata-muslo (F = 5,67; P = 0,020) con menor valor en otoño y sobre la grasa abdominal (F = 13,7; P = 0,004) con menor valor en primavera, mientras que el peso pre-faena (F = 3,96; P = 0,052) y el rendimiento (F = 3,38, P = 0,070) presentaron una tendencia cercana al significado estadístico, a ser menor y mayor respectivamente, en primavera.
La intensidad de selección aplicada durante años a las poblaciones ganaderas ha generado individuos de alta performance con una base genética mucho más estrecha que sus antepasados salvajes siendo más susceptibles ante situaciones ambientales extremas. Dado que los animales no pueden ser separados de su ambiente, ni su producción considerada en forma aislada del mismo, la productividad de las poblaciones es el resultado de una interacción genotipo x ambiente x manejo 18. La trascendencia de la interacción Genotipo x Ambiente en la cría animal fue detalladamente discutida por algunos autores 19,20,21. Desde el punto de vista estadístico existe interacción entre el genotipo y el ambiente, cuando no se verifica la suposición de que una diferencia específica de ambiente tiene el mismo efecto en todos los genotipos. Dicha respuesta diferencial puede no circunscribirse sólo al valor promedio de alguna variable respuesta sino también a la variabilidad observable en la misma. En términos genéticos podría considerarse que un determinado carácter evaluado en ambientes diferentes puede ser interpretado como caracteres diferentes, debido a que los genes que intervienen en su expresión, podrían variar en los diferentes ambientes 22. Se ha documentado el impacto negativo del stress calórico sobre la tasa de crecimiento y el consumo de alimento en diferentes poblaciones comerciales de aves para carne 23,24,25,26 y se han informado modificaciones de parámetros fisiológicos 27 de síntesis proteica y de grasa periférica subcutánea en esas condiciones 28.
La singularidad de la producción de pollos camperos, con acceso a parque y pleno impacto de las condiciones climáticas, implica un menor control ambiental en comparación con la cría industrial en confinamiento, por lo que la repercusión de los factores geoclimáticos potencia la importancia de eventuales interacciones genotipo-ambiente. Las temperaturas medias máximas registradas durante la crianza a parque y la depresión observada en el peso asintótico del 13,5% en el cruzamiento experimental Campero Casilda y del 12,7% en el genotipo de referencia, confirmaría el efecto detrimental del aumento de temperatura sobre el crecimiento en estos genotipos originariamente pensados para zonas templadas. La mayor velocidad de aproximación a un peso final menor (8,8% en el cruzamiento experimental y 7,4% en el genotipo de referencia) en primavera, no alcanzó a neutralizar la depresión en la tasa de crecimiento, presentando las aves de ambos genotipos criadas en esta estación, menor peso promedio a la edad de faena establecida por el protocolo respectivo (75-90 días). Los valores de conversión observados en los grupos estudiados, similares a los obtenidos en estudios previos en otros cruzamientos experimentales 29,30 son superiores a los registrados en la cría intensiva y a los informados por otros autores en poblaciones comerciales y autóctonas bajo crianza extensiva 31. El menor CMD observado en primavera puso en evidencia cierto efecto detrimental de las mayores temperaturas sobre la ingesta de alimento con mayor impacto de las condiciones climáticas en el genotipo de referencia, comportamiento que sugiere la presencia de interacción si bien sin significado estadístico, sobre dicha variable. No ocurrió lo mismo con el AMD, que evidenció un notable paralelismo en el comportamiento de las aves de uno y otro grupo ante el cambio de estación de crianza. Al relacionar ambas variables en términos de conversión de alimento en carne, el cruzamiento experimental con un desempeño ventajoso respecto del genotipo de referencia durante la crianza en otoño, demostró mayor estabilidad ante el cambio de estación, con similar relación de conversión (4 kg de alimento por kilo de peso producido) en ambas épocas y menor impacto de la disminución del consumo sobre la tasa de crecimiento. Campero INTA en cambio, requirió un 7,2% más de alimento por kilo de peso producido en otoño, comportamiento de clara trascendencia económica. En términos de los caracteres a la faena, los valores observados son los habituales en este tipo de aves, informados en estudios previos 32 e independientemente del genotipo, las aves criadas en primavera si bien presentaron menor peso corporal registraron mayor proporción de pata-muslo, depusieron menor proporción de grasa y presentaron mayor rendimiento a la faena.
CONCLUSIÓN
Campero Casilda es un cruzamiento experimental de tres vías propuesto como alternativa a la versión tradicional de Campero INTA, que presenta un mayor peso corporal en el rango de edades de faena permitido por el protocolo (75-90 días). Si bien en términos de relación de conversión, este genotipo mostró mayor estabilidad ante el cambio de estación, la cría con mayores temperaturas ambientales se tradujo en menor peso de faena, lo que determina disponer de un ave de menor peso, que exigirá un estudio de las relaciones existentes entre el costo de la alimentación y los ingresos derivados de la venta de un tipo de ave diferencial.

